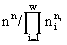 , где ni — численность в сообществе организмов вида i;
, где ni — численность в сообществе организмов вида i; 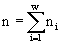 . Логарифм этого количества H
. Логарифм этого количества HБиологический факультет
Московского государственного университета им. М.В.Ломоносова
на правах рукописи
Александр Петрович Левич
Управление структурой фитопланктонных сообществ
(эксперимент и моделирование)
03.00.18 — гидробиология
Автореферат диссертации
на соискание ученой степени доктора биологических наук
Москва. 2000.
Работа выполнена на кафедре общей экологии Биологического факультета МГУ.
Научный консультант д.б.н. профессор В.Н. Максимов.
Официальные оппоненты:
чл.-корр. РАН д.б.н. профессор Г.С. Розенберг,
д.б.н. профессор Ю.Г. Симаков,
д.ф.-м.н. профессор В.В. Алексеев.
Ведущая организация: Институт глобального климата и экологии РАН.
Защита состоится 1 декабря 2000 г. в 15 часов 30 минут на заседании Диссертационного совета Д.053.05.71 на Биологическом факультете МГУ по адресу: , Москва,Воробьёвы горы,МГУ, Биологический факультет, аудитория 557.
С диссертацией можно ознакомиться в библиотеке Биологического факультета МГУ.
Отзыв в двух экземплярах просим направлять по адресу:
119899, Москва, МГУ, Биологический факультет, Диссертационный совет Д.053.05.71
Автореферат разослан ____ ноября 2000 г.
|
Ученый секретарь Диссертационного совета К.б.н. |
А.Г. Дмитриева |
Среди современных проблем водной экологии центральное место занимает проблема адекватного экологического прогноза. В приложении к экологии сообществ она могла бы быть переформулирована как проблема строгого количественного предсказания, или точнее — расчета численностей видов, образующих сообщество, как функций, аргументами которых являются факторы, определяющие жизнедеятельность организмов. Среди таких факторов одно из первых мест занимает обеспеченность особей ресурсами среды. Хорошим критерием адекватности наших расчетных схем могло бы служить умение управлять структурой сообщества с помощью внешних факторов — изменять видовой состав и доминирование, целенаправленно увеличивать или уменьшать численности выбранных групп организмов.
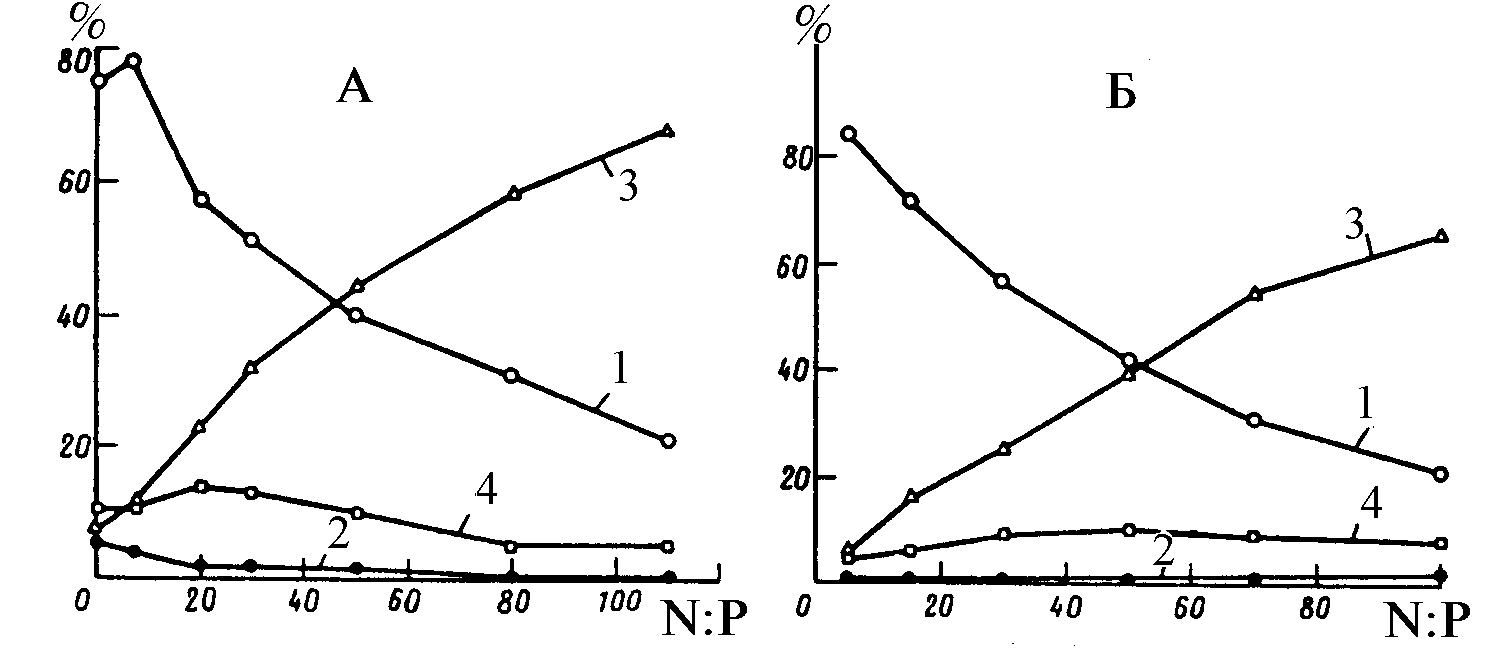
Еще в 30-е годы В.Пирсолл (Pearsall, 1930, 1932) одним из первых заметил, что различные отношения химических элементов, включая N/P, Ca+Mg/Na+K, способствуют образованию различных сообществ фитопланктона в природе. В.Смит (Smith, 1983, 1986, 1995), проанализировав связь между химическим и таксономическим составом нескольких десятков озер мира, обнаружил преобладание цианобактерий в период, когда величина атомного отношения азота к фосфору имеет значение менее 25. При отношении N/P более 25 в озерах преобладали зеленые и диатомовые микроводо-
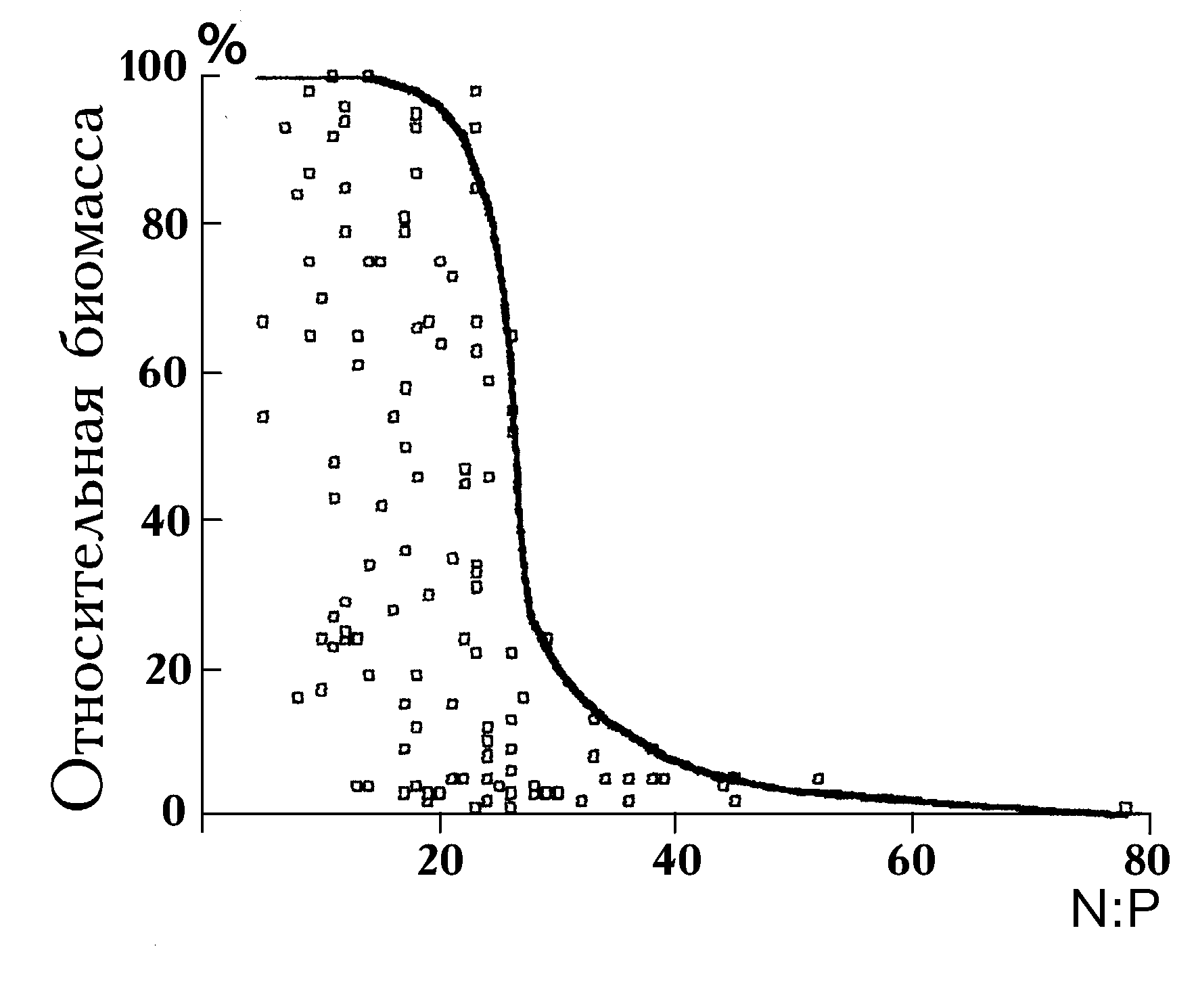
росли (рис. 1). Д.Тилман (Tilman, 1982) называет обнаруженную В.Смитом закономерность “драматическим воздействием показателя отношения азота к фосфору на таксономический состав альгоценозов озер”. Подобные эффекты и не только в связи с отношением концентраций азота и фосфора, но и для отношений, включающих кремний, микроэлементы и другие компоненты питания, наблюдали многие авторы в природных и лабораторных (проточное культивирование) условиях (Tilman, 1977; Rhee, 1978; Holm, Armstrong, 1981; Terry, 1985; Admiral, Vlugt, 1990; Shames et al., 1990; Fujimoto et
al., 1997 и др.).Отмеченные в наблюдениях гидробиологов эффекты требовали своего объяснения, интерпретаций в различных экспериментальных ситуациях, попыток прикладного применения, ответов на вопросы о роли в наблюдаемых эффектах абсолютных концентраций веществ или их отношений; о видоспецифических характеристиках клеток фитопланктона, определяющих эффекты; о выборе факторов среды, отношения которых обладают регулирующим действием на фитопланктон.
Для объяснения эффектов обычно привлекаются модели явлений. Если говорить о формальном моделировании, то наиболее распространенный в экологии путь моделирования — имитация с помощью дифференциальных уравнений. Однако на этом пути возникают трудности как принципиального, так и технического характера. Принципиальная трудность состоит в том
, что не существует систематических правил вывода самих уравнений. Процедуры их составления основываются на полуэмпирических закономерностях, правдоподобных рассуждениях, аналогиях и искусстве модельера. Технические трудности связаны с высокой размерностью задач по управлению сообществами. Для существенно многовидовых сообществ, потребляющих многочисленные ресурсы, требуется подбор сотен коэффициентов и анализ систем из десятков уравнений. (Если изучается сообщество из w групп организмов, потребляющих m ресурсов, то соответствующая система дифференциальных уравнений должна содержать, по крайней мере, w+mw+m уравнений с 2w+4mw параметрами, требующими идентификации.) Обычные приемы снижения числа переменных — их агрегирование или учет только доминирующих групп организмов — непригодны в поставленной задаче управления, поскольку в процессе управления доминирующие и малочисленные группы сменяют друг друга, а агрегация нивелирует эффекты управления. В связи с трудностями имитационного подхода управления в экологии сообществ необходима разработка альтернативных путей моделирования для анализа задач.
Цели и задачи работы. Представленная работа реализует намерение обосновать и использовать возможность регулирования состава многовидовых фитопланктонных сообществ с помощью потоков взаимонезаменимых ресурсов, потребляемых клетками. Под регулированием состава, или управлением структурой сообщества подразумевается целенаправленное изменение относительных обилий однородных интересующих исследователя групп клеток (таксонов, размерных фракций и т. п.).
Если известна эмпирическая или теоретическая зависимость численностей групп, образующих сообщество, от доступных клеткам ресурсов и каких-либо внутренних характеристик самих клеток, то поиск путей регулирования сводится к выбору значений аргументов этой зависимости, обеспечивающих необходимые численности.
Обнаружению искомой зависимости должны предшествовать, по крайней мере, два этапа исследований. Первый из них связан с отбором существенных для функционирования сообщества ресурсов
. Фитопланктонные организмы потребляют десятки взаимонезаменимых ресурсов. Однако реально на результирующие численности влияют только те из них, которых не хватает для дальнейшего развития клеток. В связи с этим возникает задача выявления ресурсов, лимитирующих рост. Вторая задача — найти видоспецифические характеристики самих клеток, ответственные за различия в отклике разных групп организмов на воздействия ресурсных факторов.Чисто эмпирическое построение искомых зависимостей представлялось заведомо бесперспективным в силу высокой размерности
, как пространства ресурсов, так и множества потенциально значимых внутриклеточных характеристик. Поэтому экспериментальному исследованию должен был предшествовать адекватный модельный анализ.Модельный анализ создавал предпосылки для экспериментального решения задач по созданию подходов к управлению альгоценозами в естественных и контролируемых лабораторных условиях.
После изучения фундаментальных закономерностей, определяющих пути регулирования состава фитопланктонных сообществ, возникли задачи по их практическому приложению в области ведения аквакультуры, деэвтрофирования “цветущих” водоемов, биологической очистки вод.
Основные
полученные результаты и их научная новизна:Основные положения диссертации, выносимые на защиту:
Публикации. По теме диссертации опубликовано 55 работ, из них четыре — монографии.
Доклады и обсуждения. За 20 лет, в течение которых велась работа над темой диссертации, состоялись многие десятки обсуждений различных аспектов работы на многочисленных семинарах, конференциях, школах и симпозиумах. Перечислю некоторые из них. Несколько организованных в 90-х годах Ростовским ГУ школ-семинаров по математическому моделированию в проблемах рационального природопользования. Второй всесоюзный съезд паразитоценологов, состоявшийся в 1983 году в городе Киеве. Школы и конференции по теоретической биологии, организованные в 80-х и 90-х годах Комиссией по теории биологических систем Министерства образования СССР и РФ. Семинар по информационным методам моделирования ВИНИТИ в 1985 г. Семинар кафедры алгебры и анализа факультета математики и механики Ленинградского ГУ в 1986 г. Семинар кафедры вычислительных методов факультета ВМиК МГУ в 1990 г. Несколько Международных конференций “Математика. Компьютер. Образование”, проводимых в последнее десятилетие кафедрой биофизики Биологического факультета МГУ. Несколько выступлений на научном семинаре кафедры гидробиологии и общей экологии Биологического факультета МГУ в 1987-89 гг. Два выступления на научном семинаре группы “Биосоляр” Географического факультета МГУ под руководством В.В.Алексеева в 1995 г. Международная конференция “Критерии самоорганизации в физических, химических и биологических системах”, прошедшая в Суздале в 1995 г. Конференция “Математическое описание ценозов”, организованная Центром системных исследований в 1996 г. Международная конференция по проблемам управления качеством окружающей среды, прошедшая в Подмосковье в 1997 г. по инициативе Правительства Москвы. Международная школа-семинар по теоретической биофизике, прошедшая в 1998 г. в Москве. Международная конференция “Анализ систем на пороге XXI века”, организованная Академией государственного управления при Президенте РФ. Несколько заседаний научного семинара Каспийского института рыбного хозяйства в 90-х годах и многие другие.
Структура диссертации. Диссертация включает введение, общую характеристику работы, пять глав, разделы с перечислением результатов работы и выводов из них, список литературы. Общий объем работы — 262 страницы. Еще на 124 страницах разместились таблицы двух приложений с числовыми данными ряда исследований. Список цитируемых публикаций насчитывает 300 работ.
Структура автореферата. Помимо сведений о диссертации автореферат включает разделы:
общая характеристика и этапы работы; объекты исследования; стадии роста культур и потребности клеток в ресурсах среды; о вариационном моделировании роста и потребления для фитопланктонных сообществ; правило лимитирующего звена; об управлении структурой фитопланктонных сообществ; концепция направленного регулирования типов “цветения” эвтрофных водоемов; подбор видов планктонных водорослей для сбалансированной утилизации многобиогенной нагрузки на водоем; формула видовой структуры и адекватность вариационной модели фитопланктонных сообществ; выводы; благодарности; список работ, опубликованных по теме диссертации.ОБЩАЯ ХАРАКТЕРИСТИКА И ЭТАПЫ РАБОТЫ
Изменение отношений количеств лимитирующих ресурсов в среде закономерным образом меняет относительные обилия видов в фитопланктонных сообществах. Идея существования такой закономерности и ее использования для управления структурой сообществ возникла при анализе
вариационной модели экологических сообществ (Левич, 1980; 1982).Модель описывает функционирование существенно многовидовых сообществ при потреблении организмами невозобновляемого запаса многочисленных взаимонезаменимых ресурсов среды. В лабораторных условиях такой ситуации соответствует накопительное, а не проточное культивирование. При проточном культивировании (в отличие от накопительного) не только возобновляется запас питательных веществ в среде, но и происходит изымание биомассы всех видов сообщества в одинаковой пропорции, обусловленной скоростью протока. Это обстоятельство существенно отличает проточное культивирование от процессов, протекающих в естественных водоемах, где элиминация биомассы почти всегда видоспецифична. В то же время накопительное культивирование может служить прообразом некоторых естественных систем, например, водоемов, в которых запасы биогенных элементов пополняются один-два раза в сезон при перемешивании обогащенных вод из-под термоклина с водными массами фотического слоя (Одум, 1986). В данном исследовании изучается период развития микроводорослей от инокуляции до остановки роста, вызванной исчерпанием одного из ресурсов, но не какими-либо иными причинами (например, не аутотоксикацией, не аллелопатическими взаимодействиями и др.). Учитывается, что рост организмов определяется не только запасами ресурсов в среде, но и их внутриклеточными их запасами. А именно, подразумевается один из физиологических механизмов влияния запасов на рост: деление клеток прекращается, когда внутриклеточный запас по одному из ресурсов достигает некоторого видоспецифического минимального значения (
Droop, 1973).Полученный в модели “на кончике пера” результат представлял собой (в лучшем случае) рабочую гипотезу, которой необходимо было пройти всестороннюю эмпирическую проверку
, прежде чем превратиться в научное положение и практическую методику или быть опровергнутой. Другими словами, предстояло реализовать целую исследовательскую программу по проверке исходной рабочей гипотезы. Эмпирическое исследование должно давать прямой и окончательный ответ о справедливости гипотезы, но оно представляет собой наиболее трудоемкую и длительную часть работы. Чтобы решиться на ее реализацию, желательно было получить, может быть, косвенные, но более легко достижимые подтверждения или опровержения исходной идеи. Таким целям мог служить целенаправленный тематический поиск фактов и суждений в научной литературе, а также дополнительный модельный анализ.Литературный поиск проводился по трем направлениям, соответствовавшим направлениям предстоящих экспериментальных работ: лабораторные альгоценозы, природный фитопланктон
in vitro, природный фитопланктон in situ. Результаты поиска обнадеживали — эффект управляющего воздействия отношений в среде азота к фосфору, азота к кремнию и др. на видовую структуру фитопланктона наблюдали многие исследователи.Был сделан шаг в развитии вариационной модели. В дополнение к утверждению о том, что относительные численности видов в сообществе зависят не от абсолютных концентраций ресурсов в среде, а от их отношений, была решена задача об оптимальных для каждого вида отношениях ресурсов. Оптимальность данного набора отношений для заданного вида означала, что относительная численность вида при этих отношениях ресурсов выше, чем при всех других. Строго доказана теорема оптимизации, утверждающая, что оптимальные для заданного вида отношения ресурсов равны отношениям потребностей этого вида в рассматриваемых ресурсах (Левич и др., 1993а;
Alexeyev, Levich, 1997). Теорема оптимизации позволяет посредством выбора в среде исходных отношений количеств ресурсов целенаправленно увеличивать или уменьшать в сообществе доли интересующих исследователя видов.Помимо вариационной модели гипотеза о роли отношений концентраций питательных веществ была испытана в традиционной имитационной модели процессов потребления и роста для фитопланктона (Левич, Личман, 1992). Эта описательная имитационная модель представляет собой систему дифференциальных уравнений для скоростей изменения биомассы каждого вида, для внутреннего пула каждого из ресурсов в каждой из популяций сообщества и для внешней концентрации в среде каждого из ресурсов. Учитывалась зависимость этих скоростей от внешних концентраций питательных веществ в форме закона Моно и от внутренних их концентраций в форме закона Друпа.
Скорости процессов в модели зависят от абсолютных значений концентраций ресурсов в среде, зависимости от отношений концентраций в модель не закладывались. Законы Моно, Друпа и сохранения вещества, использованные в модели, являются эмпирическими обобщениями, тем самым вся модель работает как “большая аппроксимирующая формула”, аккумулирующая наши опытные знания о процессах потребления и роста для фитопланктона. Машинные эксперименты с моделью показали, что в этих накопленных десятилетиями знаниях неявно заложены эффекты зависимости относительных обилий популяций сообщества от исходных отношений количеств ресурсов в среде. При этом оптимальные для каждого вида отношения оказались близки к отношениям минимальных клеточных квот для этого вида. Другими словами, наша рабочая гипотеза подтверждалась современными представлениями о функционировании фитопланктонных сообществ, заложенными в традиционную феноменологическую модель.Поскольку оказалось, что наша гипотеза не противоречит ни имеющимся литературным данным, ни традиционным представлениям о функционировании фитопланктона, то переход к экспериментальному этапу исследования в большой степени был оправдан. Оправдан, но не вполне возможен, так как на пути к прямым экспериментам существовали области “белых пятен” в теоретической и экспериментальной экологии сообществ. Во-первых, управляющими могут быть только лимитирующие рост факторы. Однако было неясно, как для многовидовых сообществ, нуждающихся в многочисленных ресурсах, прогнозировать, какие из ресурсов существенны для функционирования, или, другими словами, лимитируют деление клеток. Во-вторых, параметрами моделей и видоспецифическими параметрами управления являются потребности клеток в потребляемых ресурсах. Как само понятие потребностей, так и методы их измерения — мало разработанные области альгологии. Поэтому прежде, чем перейти к экспериментам по управлению структурой сообществ, оказалось необходимым разработать подходы к выявлению лимитирующих ресурсов для многовидовых сообществ, концепцию потребностей микроводорослей в ресурсах среды и методы измерения потребностей. Также было необходимо провести все необходимые измерения.
Лимитирующими считаются ресурсы, которые потребляются из среды и внутренних запасов полностью, в силу чего останавливается деление клеток микроводорослей и прекращается рост популяций сообщества. Полностью потребляются только лимитирующие ресурсы. Вопрос о выявлении лимитирующих ресурсов был решен на пути строгого доказательства в рамках вариационной модели теоремы стратификации пространства потребляемых сообществом ресурсов. Согласно этой теореме, пространство ресурсов разбивается на области, в каждой из которых лимитирующим рост является какое-либо одно из непустых подмножеств совокупности всех ресурсов (Левич и др., 1994). В каждой из таких областей численности видов зависят только от лимитирующих ресурсов. Теорема задает алгоритм расчета областей лимитирования, и для любой заданной совокупности значений начальных концентраций веществ можно указать, какие из ресурсов будут лимитирующими, а какие — нет. Принцип лимитирующего звена, следующий из теоремы стратификации, обобщает на многовидовые сообщества “принцип минимума” Ю.Либиха и “закон совокупного действия факторов” Э.Митчерлиха. Предложена и применена методика экспериментального выявления лимитирующих ресурсов на стационарной стадии накопительного культивирования.
Понятие клеточной потребности близко к широко используемому понятию клеточной квоты. Для применения к вопросам управления структурой сообществ была разработана концепция потребностей (Левич и др., 1986б; Левич, 1989) и предложена методика их измерения (Левич, Артюхова, 1991). Эта методика реализована для 18 видов водорослей и цианобактерий.
Для опытов в контролируемых условиях понадобилось использовать экспериментальные экологические объекты — лабораторные альгоценозы. Для создания лабораторных альгоценозов понадобилось ведение музея культур, монтирование термостатированных люминостатов и других объектов экспериментальной базы, отлаживание необходимых гидробиологических и гидрохимических методик. Опыты и обработка их результатов проводились в течение нескольких лет с культурами, включающими от 2-х до 10-ти видов фитопланктона. Были проведены 10 серий опытов, включавшие 150 отдельных ценозов, отличавшихся видовым составом фитопланктона или исходным составом среды. В этих опытах были изучены: динамика роста и потребления (Левич и др., 1986а; Артюхова и др., 1988), взаимодействие видов в поликультурах (Ревкова и др., 1985), конкуренция за ресурсы (Левич и др., 1986а; Ревкова, 1987), выявление лимитирующих ресурсов (Левич и др., 1993б; Левич, Личман, 1992), парциальные ресурсные потребности видов (Левич и др., 1986а), этапы идентификации и верификации использованных математических моделей (Левич и др., 1986а; 1986в; Левич, Лебедь, 1987; Левич, 1988; 1989; Левич, Личман, 1992; Замолодчиков и др., 1993), вопросы управления структурой сообществ (Левич, Булгаков, 1993).
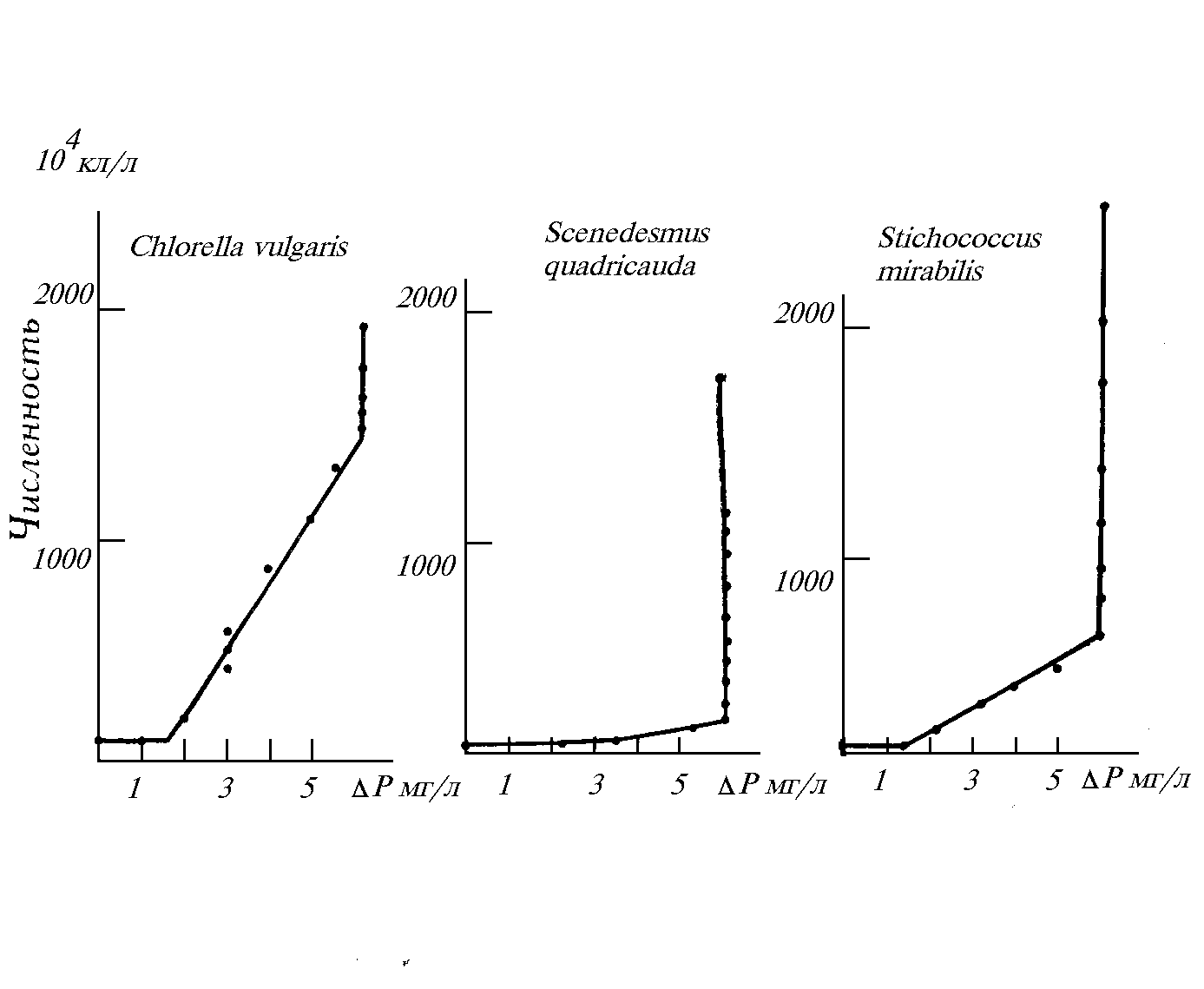
В течение нескольких сезонов проводились лабораторные опыты с природным фитопланктоном. Целью опытов была проверка гипотезы о влиянии отношения концентраций минерального азота к фосфору в исходной среде на видовую структуру микроводорослей. Сообщество фитопланктона из прудов Астраханской области насчитывало около 100 видов. Была проанализирована динамика биомассы каждого вида и отдельно — доминирующих родов, порядков, отделов и шести размерных групп. Результаты показали, что варьирование отношения
N/P в среде регулирует как видовую, так и размерную структуру природного фитопланктона in vitro (Левич и др., 1991; 1992). Изменение относительных обилий происходит как на уровне видов, так и на уровне высших таксонов (за счет реакции доминирующих в таксонах видов) и на уровне размерных групп (за счет смены доминирования в сообществе видов с разными размерами клеток). Наиболее ярко регулирование проявляется для отделов зеленых водорослей и цианобактерий: первые доминируют при высоких (20-50) атомных отношениях азота к фосфору в среде, вторые — при низких (2-5), причем доминирование сине-зеленых происходит не за счет азотфиксирующих видов. Результаты опытов in vitro не противоречат гипотезе о равенстве оптимальных отношений внешних концентраций отношениям внутриклеточных содержаний ресурсов на стационарной стадии роста.В течение пяти сезонов велись работы по испытанию влияния различных пропорций добавок азота и фосфора на таксономическую и размерную структуру фитопланктона прудов
in situ (Левич и др., 1988; 1996а;1996б; Булгаков и др., 1992; Levich, Bulgakov, 1993). Анализ таксономической структуры был проведен на уровне видов, родов, порядков и отделов водорослей. Вывод из результатов полевых испытаний: главным фактором, управляющим структурой альгофлоры в пруду, стало внесение минеральных питательных веществ в пруды в отношении, соответствующем отношению потребностей в них клеток ведущих видов из отделов зеленых, диатомовых, сине-зеленых и размерного класса 0,4-3,2 нг.“Чистые” научные исследования, как правило, открывают пути своего использования в прикладных областях человеческой деятельности. Для гидробиологических задач ведения аквакультуры, деэвтрофирования “цветущих” водоемов и биологической очистки вод были предложены методики, использующие экспериментальные и модельные результаты предшествующих фундаментальных разработок. Для обоснования практических приложений понадобился подробный литературный обзор трофической роли таксономического и размерного состава природного фитопланктона, а также роли планктона в проявлении негативных последствий избыточного биогенного обогащения поверхностных вод.
Цели ведения аквакультуры обычно требуют регулирования состава альгоценоза в направлении доминирования клеток хлорококковых, диатомовых, эвгленовых и уменьшения доли сине-зеленых, а также увеличения доли крупных видов водорослей в общей биомассе. Для рыбоводных прудов Астраханской области была предложена, апробирована и внедрена рациональная экологизированная система удобрения (Левич и др., 1988; 1996б; Булгаков и др., 1992). В эту систему был заложен целый ряд мероприятий, направленных на интенсификацию процессов продуцирования биомассы микроводорослей (увеличение частоты внесения удобрений, согласованное с динамикой их потребления, неравномерное по сезону внесение питательных веществ, резкое снижение количества вносимых фосфорных соединений, согласование доз удобрения с потребностями в них автотрофов и др.). Однако главным элементом системы, ответственным за перестройку структуры альгоценоза в прудах, являлось отношение концентраций азота и фосфора, поступающих в водоемы с минеральными удобрениями. Воздействие на фитопланктонное сообщество с помощью разработанной системы удобрения привело к повышению рыбопродуктивности опытных прудов, к заметному улучшению кислородного режима, к снижению непотребляемых водорослями остатков минерального обогащения в стоках из рыбоводных прудов.
Для использования в водоемах бытового, рекреационного, водохозяйственного, энергетического и комплексного назначения предложена концепция направленного регулирования типов “цветения”. Типичная причина эвтрофирования водоемов — увеличение нагрузки по соединениям фосфора, а типичное последствие — бурное “цветение” цианобактерий с последующим отмиранием их избыточной биомассы, выделением токсинов, нарушением кислородного режима, проявлениями гниения и т.д. С эколого-трофической точки зрения накопление в водоемах избыточной биомассы сине-зеленых при эвтрофировании есть следствие того, что большинство таксонов цианобактерий является трофическим тупиком в пищевых цепях гидробионтов. Типичный путь преодоления эвтрофирования — снижение фосфорной нагрузки на водоемы,— как правило, трудноосуществим. Предлагаемый экологический путь избавления от избыточной (вследствие фосфорного обогащения) продукции трофически неиспользуемых цианобактерий включает два шага экосистемного управления. Первый шаг — биогенное манипулирование — состоит в увеличении в воде отношения количеств азота и фосфора. Нетрадиционность метода заключается в том, что необходимое увеличение достигается не за счет снижения количества фосфора, а за счет добавления в уже эвтрофированный водоем соединений азота. Как показывают экспериментальные и модельные исследования, такая биогенная манипуляция приводит к подавлению цветения цианобактерий и доминированию хлорококковых микроводорослей. Второй шаг управления — биоманипулирование — заключается в интродукции в водоем потребителей фитопланктона — мирного зоопланктона или фитопланктоядных рыб, переводящих избыточную первичную продукцию активно потребляемых ими хлорококковых во вторичную продукцию консументов. Именно поэтому вслед за пиком “цветения” водорослей следуют пики численности зоопланктона и т. д. Таким образом, парадоксальное добавление азотных форм в обогащенный фосфором водоем приводит не к усугублению, а к ликвидации нежелательных последствий эвтрофирования (Левич, 1995б;
Levich, 1996).Биогенное управление структурой сообществ состоит в создании с помощью свойств среды необходимых исследователю соотношений в видовом составе альгоценоза. Возможна постановка и противоположной задачи управления: с помощью подбора состава сообщества осуществить полную и сбалансированную утилизацию всех биогенных компонентов содержащей водоросли среды. Подобная задача может возникнуть при решении, например, проблем биологической очистки вод. Сведения о потребностях видов в компонентах среды и расчеты областей (стратов) полного потребления (лимитирования) для произвольных подмножеств всей совокупности потребляемых сообществом ресурсов позволяют строго и однозначно решить эту задачу (Замолодчиков, Левич, 1992).
Разработка вариационной модели функционирования экологических сообществ включала несколько этапов моделирования: формулировка и интерпретации экстремального принципа (Левич, 1978; Левич, Алексеев, 1997;
Levich, 2000); расчеты с помощью функторного метода сравнения структур (Левич, 1982; 1996а; Levich, Solov’yov, 1999) функционала для вариационной задачи (обобщенной энтропии, или разнообразия сообществ); постановка и решение (Левич и др., 1994) вариационной задачи с балансовыми ограничениями по ресурсам в виде неравенств (в частности, доказательства теорем существования и единственности решения); вывод и анализ свойств формулы видовой структуры, связывающей численности видов сообщества на стационарной стадии роста с запасами лимитирующих рост ресурсов в среде и клетках и с потребностями видов в этих ресурсах (Левич, 1980; Левич и др., 1994); строгое доказательство упомянутых выше теорем стратификации, оптимизации и ряда других (Левич и др., 1993а; 1994; Alexeyev, Levich, 1997); проверка адекватности предсказаний модели (Левич и др., 1986а; 1986в; Замолодчиков и др., 1993). Формула видовой структуры позволяет: рассчитывать абсолютные и относительные численности видов как функции запасов лимитирующих ресурсов и видовых потребностей в них; рассчитывать парциальные потребления ресурсов из среды каждой из популяций; рассчитывать для нелимитирующих рост ресурсов полные их потребления из среды всем сообществом (для лимитирующих ресурсов эти потребления равны запасам); рассчитывать парциальные потребности видов в ресурсах как функции запасов ресурсов и известных численностей видов в сообществе; описывать резко убывающую форму ранговых распределений численности фитопланктона. Кроме того, вариационная модель объясняет происхождение ряда популярных в экологии индексов видового разнообразия и указывает области применимости для них.После обобщения результатов многолетней работы возникло естественное желание донести их до широкого круга исследователей. В 1995 и 1997 гг. вышли обзорные статьи о потребностях фитопланктона в ресурсах среды и об отношении азота к фосфору в среде как о самостоятельном регулирующем факторе в отечественном журнале “Успехи современной биологии” (Левич, Булгаков, 1995; 1997). В 1996 и 1997 гг. вышли в свет монографии, обобщающие различные этапы работы: А.П.Левич, Н.Г.Булгаков, Д.Г.Замолодчиков “Оптимизация структуры кормовых фитопланктонных сообществ” (под редакцией профессора В.Н.Максимова) и А.П.Левич, В.Н.Максимов, Н.Г.Булгаков “Теоретическая и экспериментальная экология фитопланктона: управление структурой и функциями сообществ”. В 1999 г. обзор наших с коллегами результатов по регуляции состава альгоценозов появился в широко цитируемом европейском журнале “
Archiv für Hydrobiologie”. В процессе рецензирования главный редактор журнала В.Ламперт предложил устроить Форум на страницах журнала для предъявления различных точек зрения на материалы нашей статьи. В августовском номере за 1999 г. вслед за нашей с Н.Г.Булгаковым статьей (Bulgakov, Levich, 1999) появились статьи известных гидробиологов С.Рейнольдса (Великобритания), В.Смита и С.Беннет (США), У.Зоммера (Германия). Опубликованные в “Archiv für Hydrobiologie” наши с коллегами результаты вызвали живой интерес гидробиологов и экологов. Аспекты вариационного моделирования были предоставлены на суд зарубежных коллег в 1997 и 1999 гг. (Alexeyev, Levich, 1997; Levich, Solov'yov, 1999) соответственно в журналах “Bulletin of Mathematical Biology” и “Cybernetics and Systems. An International Journal”. В 2000 г. в международном журнале “Ecological Modelling”, известном исключительно тщательным рецензированием, вышла статья о принципах функционирования альгоценозов и теоремах вариационного моделирования (Levich, 2000).Первичные результаты, которые могут представлять интерес независимо от их использования в настоящей работе, приведены в приложении. Это, во-первых, — размеры и массы 212 видов и форм фитопланктонных организмов, измеренные в экспериментах
in situ. Во-вторых, — это протоколы динамики численности клеток и гидрохимических измерений в 150 лабораторных экспериментах с моно- и поликультурами фитопланктона.ОСНОВНОЕ
содержание работыОбъекты исследования
Экспериментальная работа проводилась с лабораторными моно- и поли- культурами водорослей, с природным прудовым фитопланктоном in vitro и с фитопланктонными сообществами рыбоводных прудов.
Материалы 10 серий опытов с лабораторными альгоценозами были использованы для изучения взаимодействия видов, для расчетов ресурсных потребностей клеток, для идентификации и верификации моделей функционирования сообществ, для поиска путей регулирования видовой структуры сообществ и др. Полная сводка количественных результатов лабораторных опытов приведена в Приложении 3 к диссертации.
В опыте №1 поликультура состояла из 10 видов зеленых водорослей: Ankistrodesmus acicularis, A. braunii, Chlorella vulgaris, Chromochloris cinnoborina, Scenedesmus bijugatus, S. obliquus, S. quadricauda, Scotiella nivalis (Chlorophyta, Chlorococcales), Stichococcus mirabilis (Chlorophyta
, Ulothrichales), Chlamydomonas humicola (Chlorophyta, Volvocales). Поликультуры из 10 видов росли при двух уровнях освещенности — 0.15 и 4.4 Вт/м2, а монокультуры всех видов только при 4.4 Вт/м2. Культуры выращивались на одной среде с исходным содержанием азота (N) 195 мг/л, фосфора (P) — 5.2 мг/л в течение 68 суток.В опыте №2 участвовали 8 видов водорослей: Ankistrodesmus acicularis, A. falcatus, Chlorella vulgaris, Scenedesmus quadricauda, Scotiella nivalis (Chlorophyta, Chlorococcales), Stichococcus mirabilis (Chlorophyta, Ulothrichales), Anacystis nidulans (Cyanophyta) и Pleurochloris magna (Xanthophyta). Продолжительность эксперимента составила 36 суток. Культуры росли на одной среде (N — 100 мг/л, P — 6 мг/л) в трех повторностях. В двух из них исходные плотности видов в поликультуре были примерно равны исходным плотностям соответствующих монокультур. В третьей повторности начальные численности были значительно меньше, так что суммарная численность поликультуры была сравнима со средним посевным титром монокультур.
В опыте №3 количество входящих в сообщество видов было сокращено до четырех. Все они принадлежали к порядку Chlorococcales: Ankistrodesmus sp., Chlorella vulgaris, Scenedesmus quadricauda, S. obliquus. Использовали пять сред, отличавшихся содержанием фосфора: 1, 2, 3, 4 и 5 мг/л. Азота всюду добавляли 30 мг/л. Опыт продолжался 30 суток.
Опыт №4 состоял в изучении роста пяти видов хлорококковых водорослей: Ankistrodesmus sp., A. falcatus, Chlorella vulgaris, Coelastrum sp., Scenedesmus quadricauda на двух средах: среда 1: N — 11 мг/л, P — 3.5 мг/л; среда 2: N — 60 мг/л, P — 3 мг/л. Опыт продолжался 37 суток.
Два вида хлорококковых составили простейшую поликультуру в опыте №5. Scenedesmus obliquus и S. quadricauda выращивали на трех средах с одинаковым исходным содержанием фосфора 4.5 мг/л. Концентрации азота составили 11.5, 37 и 110 мг/л. Текущие пробы отбирали в течение 82 суток. В результате в конце опыта наблюдали очень высокую плотность клеток, которая вызывала лимитирование по свету.
В опыте №6 поликультура состояла только из представителей порядка Chlorococcales: Ankistrodesmus sp., Chlorella vulgaris, Scenedesmus quadricauda. Испытывали два варианта сочетания добавок азота и фосфора. Концентрации фосфора в обоих вариантах равнялись 4.5 мг/л, азота — 30 и 60 мг/л. Длительность опыта составила 45 суток.
Четыре ранее изучавшихся вида хлорококковых были исследованы в опыте №7. Это Ankistrodesmus falcatus, Chlorella vulgaris, Scenedesmus obliquus и S. quadricauda. Культивирование проходило на трех средах с разным содержанием фосфора (среда 1 — 0.5 мг/л; среда 2 — 1 мг/л; среда 3 — 1.5 мг/л) и одинаковым содержанием азота — 30 мг/л. Было создано несколько альгоценозов. Поликультура из всех четырех видов росла на всех средах. Помимо этого, поликультуры из трех видов (четыре возможных сочетания) культивировали на средах с содержанием фосфора 1 мг/л. На 14-е сутки опыта, когда в среде оставалось около 20 мг/л азота, во все колбы было добавлено по 40 мг/л этого элемента, и опыт продолжался еще 16 суток.
Поликультуру из трех видов хлорококковых Ankistrodesmus acicularis, Chlorella ellipsoidea, Scenedesmus quadricauda испытывали в опыте №8 при начальной концентрации фосфора 3 мг/л; концентрации азота имели три уровня: 6, 12 и 19 мг/л. Кроме трехвидовой, изучали также три двухвидовые поликультуры на всех трех средах. Длительность опыта составила 28 суток.
В опыте №9 сравнивали рост в поликультуре двух видов из порядка хлорококковых Ankistrodesmus falcatus и Scenedesmus quadricauda на средах, значительно различающихся соотношением исходных концентраций биогенных элементов: среда 1: N — 14 мг/л; P — 3.1 мг/л; N/P = 4.5; среда 2: N — 34 мг/л; P — 0.6 мг/л; N/P = 57; среда 3: N — 34 мг/л; P — 0.1 мг/л; N/P = 340; среда 4: N — 4 мг/л; P — 3.1 мг/л; N/P = 1.3. Опыт продолжался 56 суток и в конце его было обнаружено лимитирование светом.
В опыте №10 в состав сообщества были включены Ankistrodesmus falcatus, Scenedesmus quadricauda (Chlorophyta, Chlorococcales) и Anabaena variabilis (Cyanophyta). Как и в предыдущем эксперименте, были выбраны четыре среды с разным соотношением азота и фосфора: среда 1: N — 0.05 мг/л; P — 0.01 мг/л; N/P = 5; среда 2: N — 0.25 мг/л; P — 0.01 мг/л; N/P = 25; среда 3: N — 0.75 мг/л; P — 0.01 мг/л; N/P = 75; среда 4: N — 2 мг/л; P — 0.01 мг/л; N/P = 200. Низкие абсолютные концентрации элементов обусловили быстрое наступление стационарной фазы (на 24-е сутки). Трехвидовая поликультура росла на всех четырех средах, а три поликультуры с попарным сочетанием видов — на двух средах (N — 0.05 и 0.75 мг/л).
Эксперименты с природным фитопланктоном
in vitro проводились в контролируемых условиях. Воду из рыбоводного пруда помещали в шесть аквариумов объемом 20 л, в которые затем добавляли суперфосфат и аммиачную селитру в разных количественных сочетаниях. С учетом природного фона в двух аквариумах отношение N/P составило 5, в двух — 12, в двух — 16. Исходные биомассы каждого вида во всех аквариумах были одинаковы. Чтобы исключить эффект выедания зоопланктоном, опытную воду перед внесением добавок пропускали через ячеистую сеть с соответствующим размером ячей и оставляли на двое суток в темноте. В одном из аквариумов исходная концентрация биогенных элементов была равна фоновой. В течение опыта все аквариумы содержали на открытом воздухе. Для некоторых абсолютных концентраций и соотношений существовали повторности, по которым для всех фитопланктонных показателей проводили усреднения. С помощью микроскопирования определяли численность и одновременно массы клеток фитопланктона (путем измерения индивидуальных размеров). Полученная биомасса служила основным функциональным показателем различных систематических групп. Наблюдения за динамикой роста водорослей вели в течение 14 суток. Схема второго опыта с более широким набором исходных отношений биогенных элементов (отношение N/P составляло 2, 5, 20, 50 и 100) не имела принципиальных отличий за исключением того, что постановку опытов осуществляли в двухлитровых колбах.
Опыты с естественным фитопланктоном
in situ в течении 5 лет велись в семи экспериментальных нагульных рыбоводных прудах Астраханской области. Площади прудов составляли 0,25 – 3,5 га, глубина — около 1,5 метров. В опытные пруды с начала апреля по начало сентября вносили два раза в неделю азотные и фосфорные удобрения в виде соответственно аммиачной селитры и простого суперфосфата. В апреле-мае отношение количеств азота и фосфора в удобрениях в расчете на элемент равнялось 4, а на протяжении остальной части сезона оно менялось от 25 до 50. В первые два внесения удобрений, непосредственно перед зарыблением, в пруды добавляли повышенные дозы биогеных элементов для обеспечения возрастающих потребностей микроводорослей в период лаг-фазы. Высокое отношение азота к фосфору в удобрениях в течение большей части всех сезонов достигалось за счет сезонных доз азотных удобрений, увеличенных в 2-3 раза по сравнению с контролем, и уменьшенных примерно в 3 раза доз фосфорных удобрений. Обязательным элементом удобрения экспериментальных прудов была также неравномерная по сезону динамика внесения минеральных веществ в соответствии с меняющимися в течение года пищевыми потребностями фитопланктона, зоопланктона и растительноядных рыб. В контрольные пруды те же минеральные удобрения вносились раз в 10-15 дней с отношением азота к фосфору, равным 4, причем абсолютные дозы вносимых веществ в течение сезона были постоянными.С апреля по сентябрь во всех прудах раз в 10 дней отбирались пробы фитопланктона и проводились гидрохимические измерения, в частности, устанавливались концентрации в воде минеральных форм азота и фосфора. Путем микроскопирования определялась численность отдельных видов фитопланктона. Параллельно измерялись индивидуальные размеры клеток, затем с помощью размеров вычислялись биомассы популяций (данные о размерах и биомассах клеток фитопланктона, полученные в этих экспериментах, помещены в Приложении 2 к диссертации). Анализ таксономической структуры фитопланктонного сообщества велся на уровне видов, родов, порядков и отделов микроводорослей. Для исследования влияния экспериментального удобрения на размерную структуру сообщества все фитопланктонные организмы классифицировали по размерам особей в единицах массы.
Стадии роста культур и потребности клеток в ресурсах среды
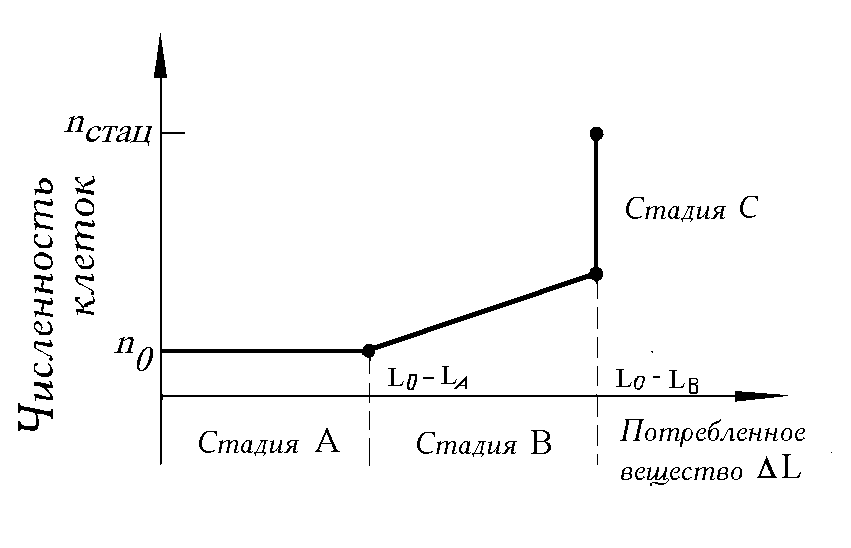
В многочисленных лабораторных экспериментах, проведенных автором и его коллегами (Левич и др., 1986б) с накопительными культурами фитопланктона, установлено, что процесс роста биомассы (численности) клеточных популяций, обусловленный потреблением биогенных элементов, закономерно складывается из трех стадий (рис. 2):
стадия A — поглощение веществ клетками и накопление биогенов без увеличения численности клеток;
стадия B — деление клеток при одновременном потреблении вещества из среды;
стадия C — деление клеток без потребления веществ из среды, т. е. за счет внутренних запасов питательного вещества (см. также Droop, 1968; Caperon, 1968;
Смирнов, 1982; Болсуновский, 1999).Данные стадии, называемые составными частями процесса “потребление-рост”, могут быть перекрывающимися и редуцированными. Стадии могут лишь частично совпадать с фазами роста, выявленными по кривым роста. Т.е. стадия A является составной частью лаг - фазы (на начальном этапе лагфазы нет ни потребления, ни роста), а стадии B и C входят в фазу экспоненциального роста.
При экспериментальном изучении роста популяции микроорганизмов стадия В наблюдается как при периодическом (накопительном), так и при проточном (непрерывном) культивировании, когда деление клеток происходит за счет веществ, поступающих в культиватор постоянно со свежей средой. Стадия С, как правило, составляет наибольшую по длительности фазу при накопительном культивировании. В природных водоемах рост популяции происходит в режимах, промежуточных между непрерывным и накопительным — приток биогенов может быть неоднократным в течение сезона, но завершается сезон, как правило, при отсутствии в среде веществ, лимитирующих рост.
На рис. 3 в качестве иллюстраций приведены некоторые экспериментальные варианты идеализированной зависимости “потребление-рост” для ряда изученных видов в условиях лимитирования по фосфору.
Под потребностями клеток фитопланктона в ресурсах среды понимается количество ресурса, поглощенное из среды одной клеткой и находящееся в клетке в процессе ее функционирования. Потребности клеток меняются от стадии к стадии роста и видоспецифичны. Измерение потребностей возможно гидробиологическим методом (
Dauta et al. 1982; Ревкова, 1987; Левич, Артюхова, 1991), при котором измеряются убыль биогенного элемента из среды и прирост численности клеток за то же время. При вычислении потребностей следует учитывать и запасы вещества в клетках к началу эксперимента. В таблице 1 сведены величины потребностей в азоте и фосфоре для микроводорослей, изучавшихся в лаборатории при накопительном режиме культивирования. В дальнейшем подходы к оценкам клеточных потребностей были развиты Н.А.Смирновым и Н.В.Ревковой (Ревкова, 1998).
Таблица 1
Потребности в азоте и фосфоре (10-9 мг/кл) для разных водорослей по результатам различных экспериментальных серий и литературным данным (отмечены звездочками). Литературные источники: * — Dauta, 1982b; ** — Muller, 1972; *** — Grover, 1989
|
Вид |
Потребность в азоте |
Потребность в фосфоре |
||||
|
Водоросли |
К концу стадии А |
На стадии В |
К концу стадии С |
К концу стадии А |
На стадии В |
К концу стадии С |
|
Сhlorella ellipsoidea |
1.5 -56.5 |
1.6± 0.19 |
1.5± 0.19 |
1± 4 |
0.25± 0.07 |
0.04± 0.01 |
|
Ch. sp. |
2 -15 |
2.8± 0.5 |
< 0.6 |
0.6 -2.6 |
1.1 -0.16 |
< 0.02 |
|
Scenedesmus quadricauda |
5 -6 |
3.3± 0.36 |
< 1.2 |
0.2 -0.5 |
1.2 -0.3 |
< 0.02 |
|
S. obliquus |
- |
1.9± 0.2 |
0.46± 0.04 |
1 -3.8 |
0.2± 0.04 |
0.025 |
|
Ankistrodesmus falcatus |
- |
3.0± 0.6 |
0.4± 0.08 |
1.25 |
0.2± 0.1 |
< 0.04 |
|
Anacystis nidulans |
- |
0.1± 0.03 |
0.05± 0.02 |
0.01 -0.2 |
0.02± 0.01 |
0.0025± 0.0003 |
|
Anabaena variabilis |
0.4-23 |
2.1± 0.27 |
1.95± 0.63 |
0.2 -0.5 |
0.2± 0.1 |
0.09± 0.04 |
|
Ankistrodesmus acicularis |
13.7 4 |
0.33 0.2 |
||||
|
Ankistrodesmus braunii |
5 |
0.2 |
12.89 5.5 |
0.11 0.16 |
||
|
Ankistrodesmus sp. |
18 |
4.17 4.3 4.9 |
1.57 |
4.76 2.7 3.5 |
0.89 0.53 0.5 0.74 |
0.12 |
|
Chlamydomonas gyrus |
8.5 |
0.75 |
1.4 |
2.7 |
0.08 |
|
|
Chlamydomonas humicola |
0.76 |
1.2 |
||||
|
Chlorella vulgaris |
|
2.2 3.9
2.9 |
0.9 0.83 |
2.41 1.5 1.16
|
0.19 0.4 0.6 0.29 0.48
0.8 |
0.4 0.3 0.05
0.13 |
|
Chromochloris cinnoborina |
2.92 |
3.08 |
3.1 |
|||
|
Pleurochloris magna |
2.9 |
0.5 |
1.3 |
|||
|
Scenedesmus bijugatus |
10 |
0.8 |
5.69 |
1.01 1.2 |
||
|
Scenedesmus *quadricauda ** *** |
0.23 0.14 0.12 |
|||||
|
Scotiella nivalis |
36.12 |
0.88 |
||||
|
Stichococcus mirabilis |
1.1 1.1 |
0.06 0.08 |
10.93 0.17 7.1 |
0.57 0.03 0.9 |
||
О вариационном моделировании роста и потребления для фитопланктонных сообществ
Для анализа существенно многовидовых сообществ в условиях потребления многочисленных ресурсов была разработана вариационная модель потребления и роста для фитопланктона. Эта модель позволяет не угадывать, а выводить основные уравнения экологии сообществ. Модель сводится к анализу m алгебраических, а не дифференциальных уравнений с mw видоспецифическискими параметрами.
Экологическое сообщество моделируется с помощью структуры множеств с разбиениями. Элементы множества — особи, классы разбиения — популяции клеток. Переходы сообщества из одних состояний в другие описываются соответствиями между множествами с разбиениями. Пусть сообщество фитопланктона состоит из w видов и потребляет m взаимонезаменимых ресурсов. Пусть в сообществе допустимы деление клеток и их гибель, но недопустимы поглощение одних организмов другими и интродукция особей извне. Таким требованиям удовлетворяют инъективные, сюръективные, не всюду определенные и не функциональные соответствия. В модели принят экстремальный принцип: в реальности осуществляется состояние, структура которого экстремальна. Согласно функторному методу сравнения математических структур (Левич, 1982), экстремальной структуре соответствует экстремальное количество допустимых структурой преобразований. Для принятых выше допущений удельное количество преобразований есть , где ni — численность в сообществе организмов вида i; . Логарифм этого количества H
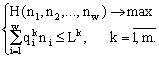
Здесь Lk — запас ресурса k в среде и внутри клеток; ![]() — потребность организма вида i в ресурсе k. Балансовые ограничения имеют форму неравенств, а не равенств, поскольку обычно известно, какой набор ресурсов необходим сообществу, но неизвестно, какие из этих ресурсов потребляются полностью.
— потребность организма вида i в ресурсе k. Балансовые ограничения имеют форму неравенств, а не равенств, поскольку обычно известно, какой набор ресурсов необходим сообществу, но неизвестно, какие из этих ресурсов потребляются полностью.
Для указанной выше вариационной задачи доказаны существование и единственность решения; найдено это решение — формула видовой структуры; доказана теорема стратификации, задающая строгий алгоритм выявления лимитирующих развитие сообщества ресурсов; доказана теорема оптимизации, позволяющая выбирать отношение ресурсов в среде, обеспечивающее наибольшие значения относительных обилий в сообществе для заданных видов; доказана теорема Гиббса, согласно которой требование максимума видового разнообразия при ограничениях сверху на потребляемые ресурсы равносильно требованию минимума потребления лимитирующих ресурсов при ограничениях
снизу на структурные параметры сообщества.Вариационная модель (как, впрочем, и описанная в разделе 3.5 модель имитационная) не вскрывает физиологические механизмы влияния начальных концентраций ресурсов на конечные относительные обилия. Вариационная модель основана на телеологическом экстремальном принципе — постулате о стремлении структуры сообщества к максимальному видовому разнообразию, допустимому имеющимся запасом ресурсов. Связь между отношением исходных концентраций ресурсов и конечными относительными численностями — формальный результат применения исходного постулата. В объяснении результатов конечными целями (телеологии) — и сила, и слабость вариационных моделей. Слабость очевидна — отсутствие механизмов, объясняющих результаты. Сила — в возможности предсказаний даже в тех случаях, когда имеющийся уровень развития знаний не позволяет заложить в модель конкретные механизмы, ведущие от причин к следствиям. Модель отличает еще то обстоятельство, что она не только адекватно предсказывает исход конкуренции, но и указывает физиологические параметры видов — их потребность в лимитирующих ресурсах, которые определяют предсказанный исход.
Правило лимитирующего звена
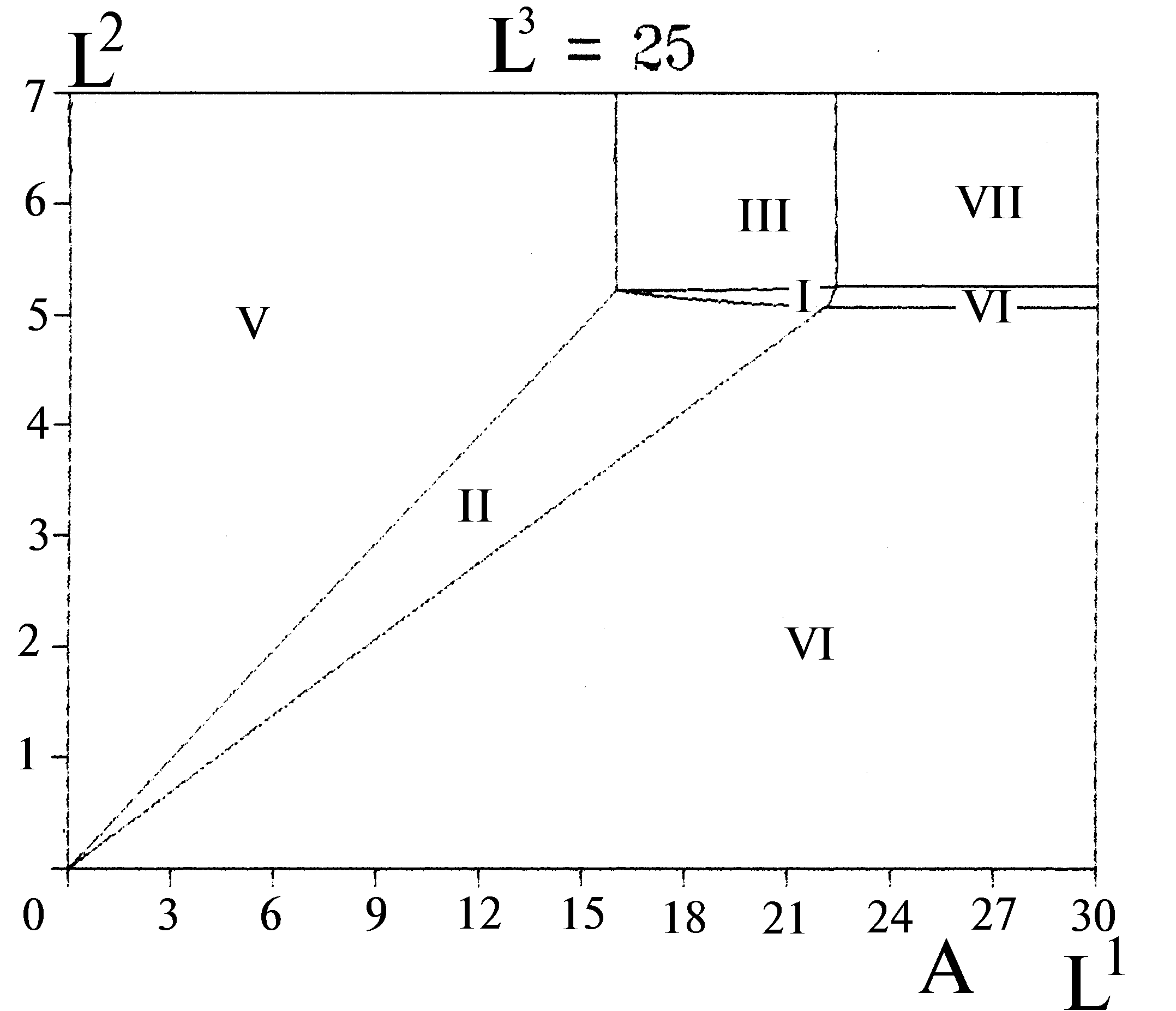
Согласно вариационной модели пространство ресурсов среды распадается на области, в каждой из которых лимитирующими рост сообщества оказываются ресурсы из некоторого подмножества совокупности всех потребляемых сообществом ресурсов. Для любого непустого подмножества ресурсов найдется единственная такая область лимитирования. В любой части пространства ресурсов лимитирующим оказывается хотя бы один ресурс. Если сообщество потребляет
m ресурсов, то в пространстве ресурсов образуется 2m-1 областей лимитирования. Лимитирующими рост считаются ресурсы, исчерпание которых приводит к остановке роста (а добавление в среду — приводит к его возобновлению). При накопительном культивировании лимитирующие вещества (и только они) потребляются из среды полностью, а балансовые неравенства в формулировке вариационной задачи превращаются в равенства. Указанные утверждения представляют собой формулировки “правила лимитирующего звена”, или строгой теоремы стратификации. Теорема задает однозначный алгоритм расчета областей лимитирования (стратов) для заданного в сообществе набора квот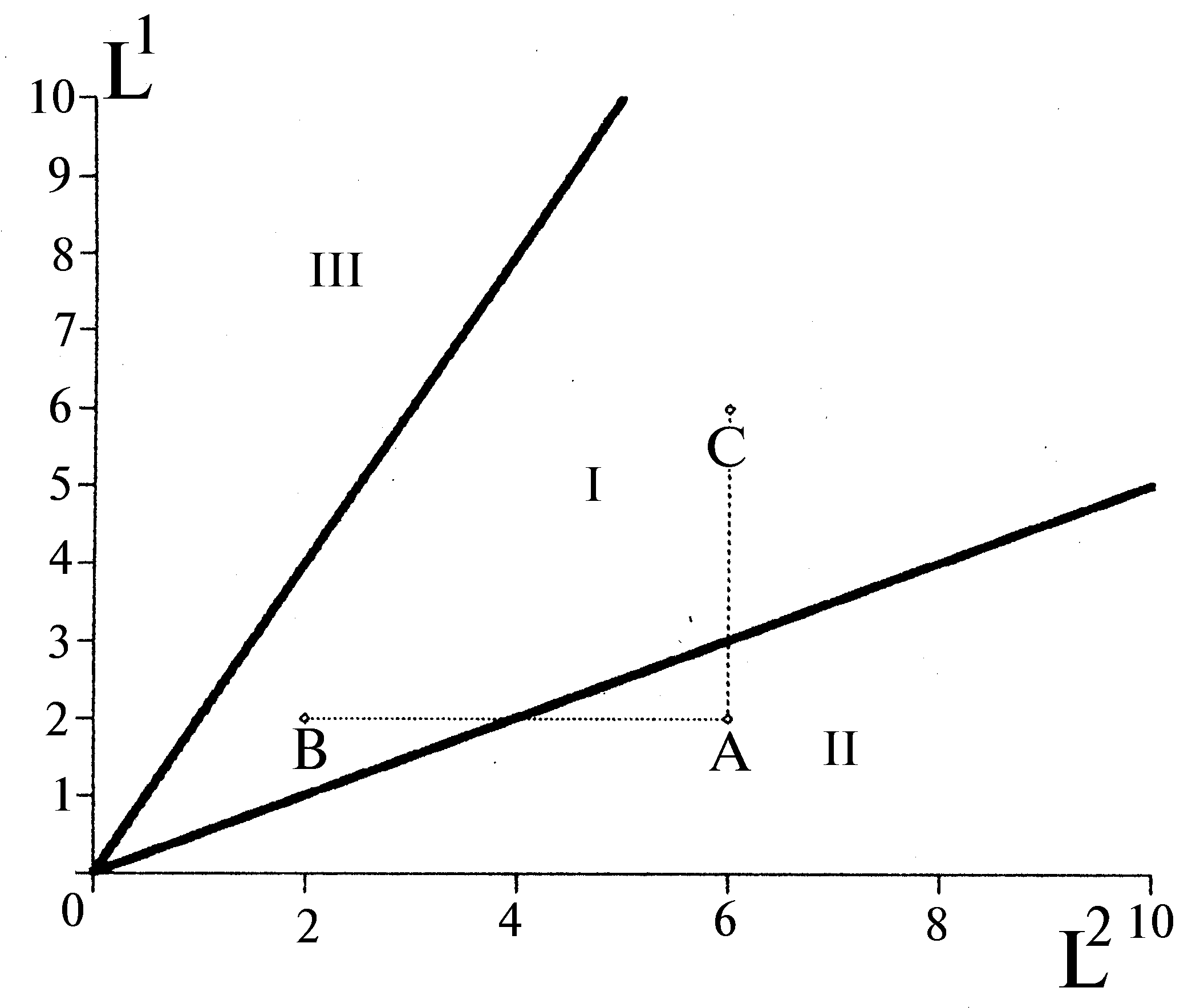
Для монокультур правило лимитирующего звена совпадает с принципом минимума Либиха (
Liebig, 1840), а именно приПо алгоритмам теоремы стратификации для заданных запасов ресурсов можно строго указать, какие из них будут ограничивать рост сообщества на стационарной стадии, а какие — нет.
Примеры расчетов приведены в диссертации и соответствующих публикациях.Опишу прямой эксперимент по проверке правила лимитирующего звена. Для альгоценоза с известными потребностями видов ![]() (i=1,…,w; k=1,…,m)
(i=1,…,w; k=1,…,m)
Подобные эксперименты были проведены для двух лимитирующих ресурсов — азота и фосфора в нескольких опытах с лабораторными альгоценозами. Потребности видов
qiN и qiP определялись в опытах с монокультурами. Для каждого из трех стратов в пространстве двух ресурсов (рис. 3) на стационарной стадии роста производились пересевы поликультур в четыре новые колбы. В первую из них вносили дистиллированную воду с добавкой азота, во вторую — с добавкой фосфора, в третью колбу — с добавками и азота, и фосфора, в четвертую вносили только дистиллированную воду (чтобы проверить, не был ли рост поликультур в исходном опыте ограничен недостатком объема среды и связанным с ним самозатенением). Для каждого из стратов рост должен был возобновляться в одной из трех колб с добавками биогенных элементов из данного страта, что и наблюдалось в подавляющем большинстве случаев.
Об управлении структурой фитопланктонных сообществ
В рамках вариационной модели доказано, что относительные обилия видов в сообществе зависят только от отношений лимитирующих рост клеток ресурсов среды. Также доказано, что относительное обилие некоторого вида принимает свое наибольшее значение при отношениях ресурсов в среде, равном отношениям потребностей в этих ресурсах у данного вида.
Феноменологическая верификация.
Существует ряд хорошо установленных феноменологических зависимостей, отражающих эмпирические представления о росте и потреблении фитопланктона. К ним относятся, в частности, закон Моно (Monod, 1942) для поглощения ресурсов из среды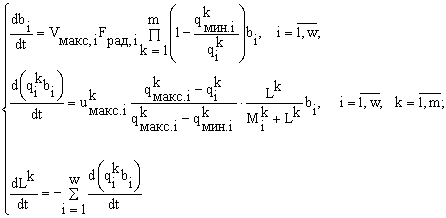
для w
=3 и 4; m=2 (Левич, Личман, 1992). Помимо уже введенных обозначений, здесь: bi — биомасса вида i; Vмакс. и uмакс. — наибольшие скорости соответственно роста и поглощения, аПосле калибровки модели по данным экспериментов с лабораторным альгоценозом были проведены три машинных эксперимента соответственно с 45, 240 и 17 комбинациями начальных концентраций азота и фосфора. Зависимость видовых обилий от отношения
N/P не закладывалась в модель. Результаты имитаций подтверждают гипотезу о зависимости относительных численностей от отношения N/P и о близости оптимального отношения N/P к отношению соответствующих откалиброванных минимальных квот для каждого из видов (табл. 2, рис. 6).Экспериментальная регуляция состава альгоценоза с помощью отношения ресурсов в среде. Собственные эксперименты проводились с лабораторными поликультурами и с сообществами естественного фитопланктона in vitro и in situ.
Проанализируем несколько экспериментов с лабораторными альгоценозами. В одном из опытов (опыт №4) участвовали четыре вида хлорококковых водорослей Scenedesmus quadricauda (Turp.) Brеb., Сhlorella vulgaris Beyer, Ankistrodesmus falcatus (Corda) Ralfs и Ankistrodesmus sp., поликультура из которых росла на двух средах со следующими начальными концентрациями биогенных элементов: среда 1 — 11 мг/л азота и 3 мг/л фосфора (N/P
= 3.5); среда 2 — 50 мг/л азота и 2.5 мг/л фосфора (N/Р = 20).
Таблица 2
Зависимость относительных численностей видов в сообществе от отношения N
/P в среде при имитационном моделировании|
Процент вида в сообществе |
||||
|
N : P |
S.q. |
S.о. |
A.f. |
C.v. |
|
0.07-5 |
75-80 |
3-6 |
6-8 |
5-10 |
|
5-10 |
70-85 |
1-6 |
6-15 |
5-15 |
|
10-20 |
60-75 |
0,6-4 |
10-20 |
6-17 |
|
20-30 |
40-60 |
0,5-3 |
20-40 |
10-17 |
|
30-60 |
35-50 |
1-3 |
37-55 |
7-15 |
|
60-80 |
30-40 |
1,5-2 |
50-60 |
7-9 |
|
80-220 |
10-25 |
1-2 |
60-85 |
4-8 |
Сокращения: S.q. — S. quadricauda, S.o. — S. obliquus, A.f. — A. falcatus, C.v. — Ch. vulgaris, N — азот, P — фосфор.
Искусственное сообщество другого опыта (опыт №9) состояло только из двух видов хлорококковых S. quadricauda и A. falcatus, но при большем наборе соотношений азота и фосфора: среда 1 — 4 мг/л азота и 3.1 мг/л фосфора (N/Р
= 1.3); среда 2 — 14 мг/л азота и 3.1 мг/л фосфора (N/Р = 4.5); среда 3 — 34 мг/л азота и 0.6 мг/л фосфора (N/Р = 57).Продолжительность опыта №4 составила 37 суток. На обеих средах с учетом роста на запасах и субстратах среды в сообществе доминировал вид S. quadricauda. Однако, если при отношении азота к фосфору, равном 3.5, это доминирование было не столь безусловным (44% от общей численности), то при увеличении отношения до 20 культура S. quadricauda практически вытеснила три других вида из сообщества (рис. 7А). Относительная численность последних в сумме составила 17%. Абсолютная численность, например, Ch. vulgaris на среде с высоким отношением N/P в 6 раз меньше, чем на среде с низким отношением. При N/P
= 3.5 стационарной фазы роста достигли только популяции S. quadricauda и A. falcatus. При отношении, равном 20, наоборот, вид S. quadricauda был единственным, продолжавшим расти в момент окончания эксперимента. Поэтому доведение поликультуры до полной остановки деления скорей всего привело бы к усилению обнаруженной тенденции.Сравнение отношений конечных численностей видов, выросших только на азоте и фосфоре среды, показало, что при переходе от низкого отношения N:Р к высокому меняется доминирующий вид сообщества. Если в первой серии опытов при N/P
= 3.5 абсолютно доминирует Ch. vulgaris (100%), то при N/P = 20 около 93% общей биомассы составляет биомасса S. quadricauda (рис. 7Б).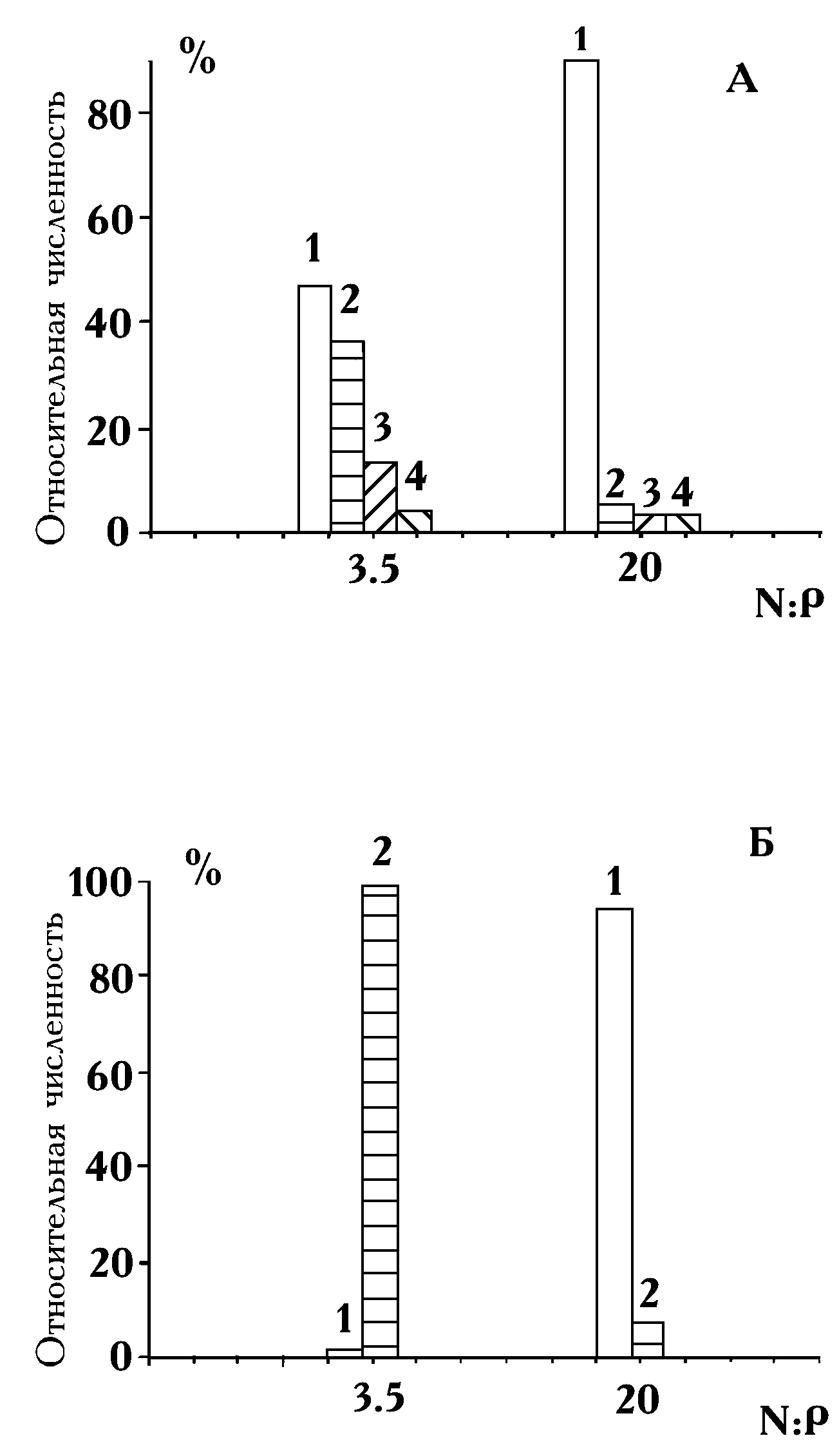
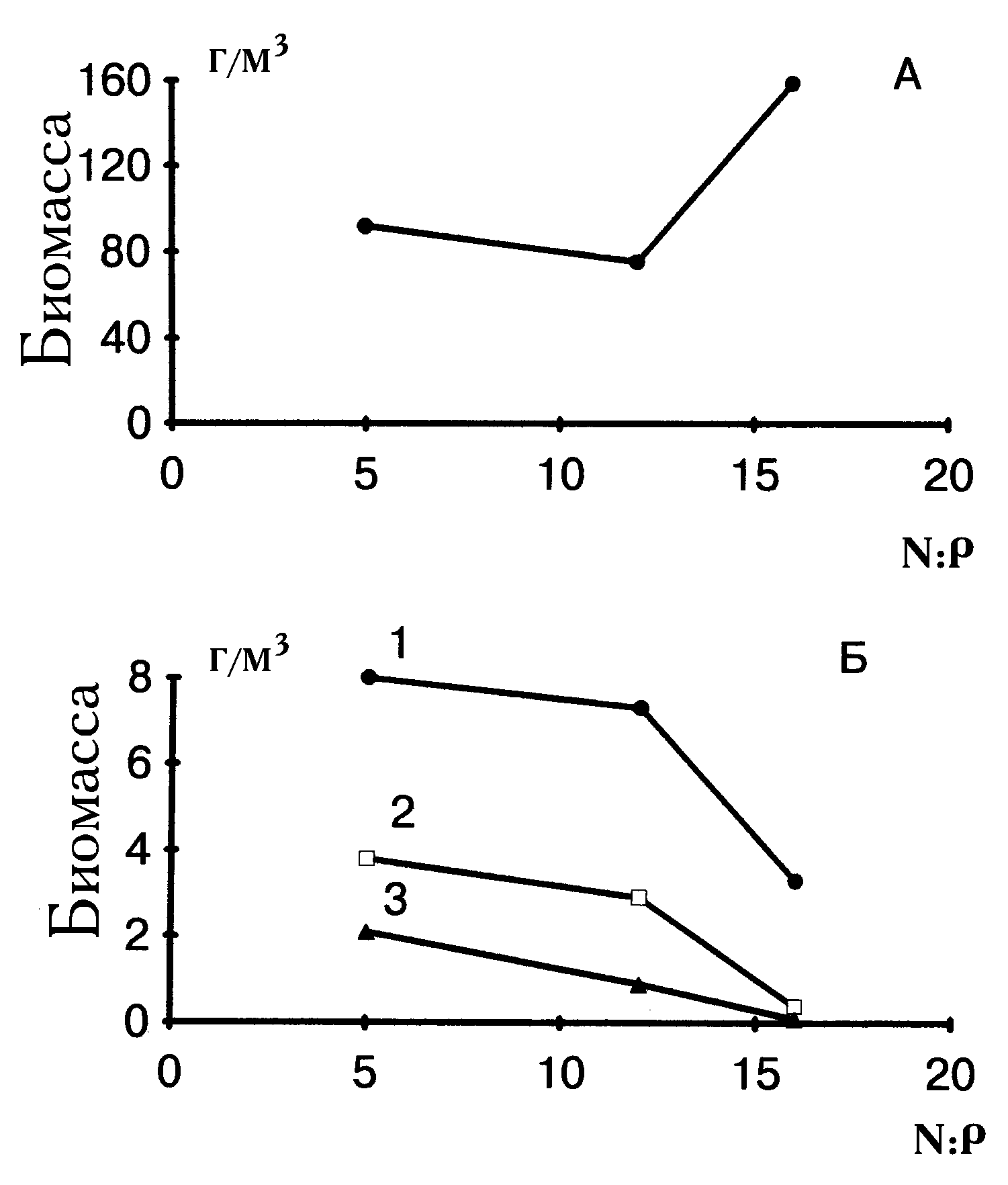 Результаты исследования роста двух видов в течение двух месяцев в опыте №9 проиллюстрированы на рис. 8. При посеве инокулятов культур во всех средах поликультура состояла на 20% из клеток S. quadricauda и на 80% — из A. falcatus. По мере увеличения отношения азота к фосфору меняется и соотношение конечных обилий двух видов. S. quadricauda увеличивает свое видовое обилие и в лимитированных азотом средах (отношения 1.3 и 4.5), и в среде, ограниченной по фосфору (отношение 57).
Результаты исследования роста двух видов в течение двух месяцев в опыте №9 проиллюстрированы на рис. 8. При посеве инокулятов культур во всех средах поликультура состояла на 20% из клеток S. quadricauda и на 80% — из A. falcatus. По мере увеличения отношения азота к фосфору меняется и соотношение конечных обилий двух видов. S. quadricauda увеличивает свое видовое обилие и в лимитированных азотом средах (отношения 1.3 и 4.5), и в среде, ограниченной по фосфору (отношение 57).
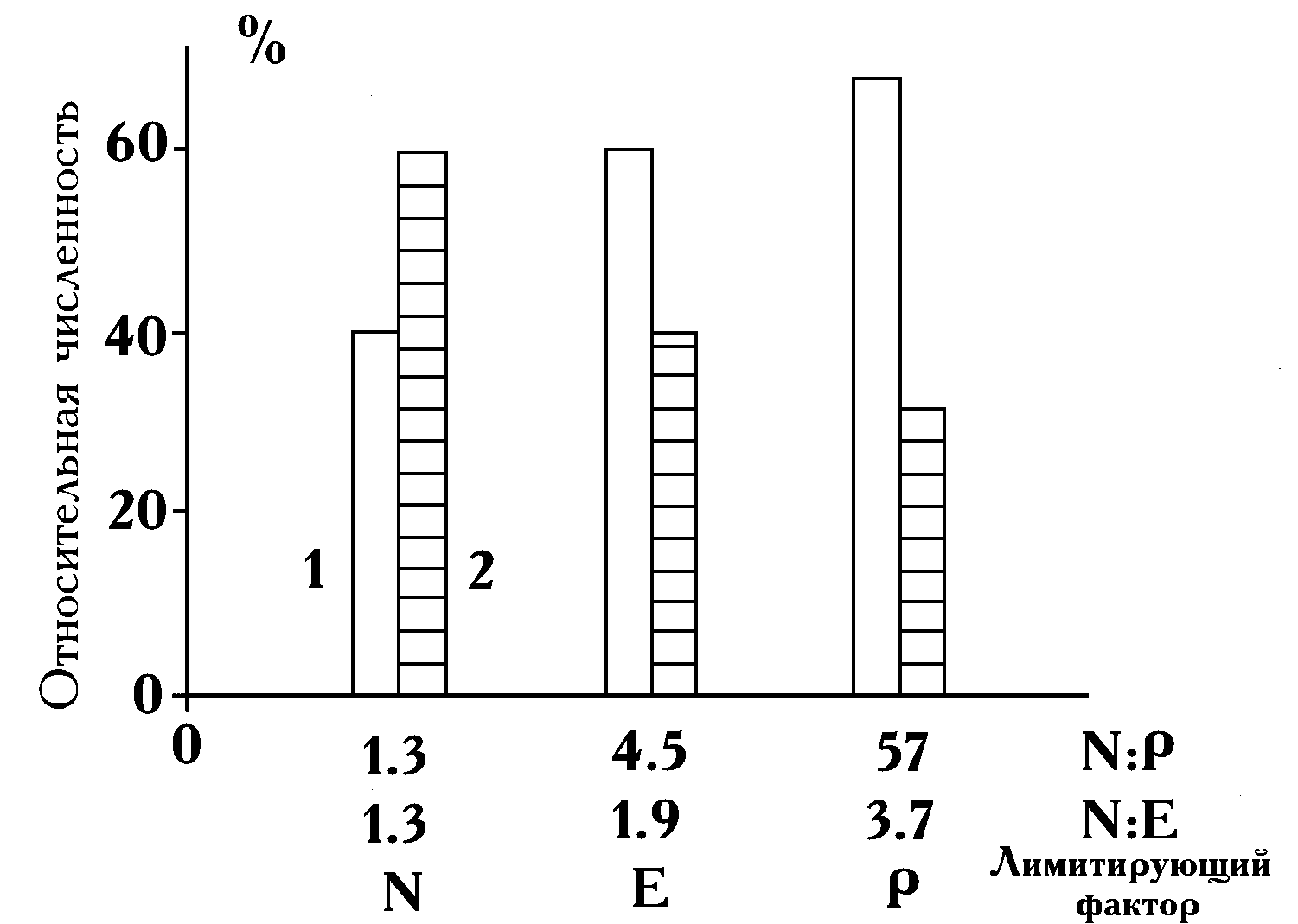
(
В диссертации содержится также анализ ситуаций с ограничениями по световой энергии E.) Изменение его относительной численности происходит при отношениях 1.3, 4.5 и 57 — соответственно от 40 до 60 и 68%. В этой серии рост культур на запасах был невелик по сравнению с общим приростом, поэтому соответствующие поправки не внесли существенных изменений в распределение биомасс видов на всех трех средах. Анализ ряда других лабораторных экспериментов содержится в диссертации.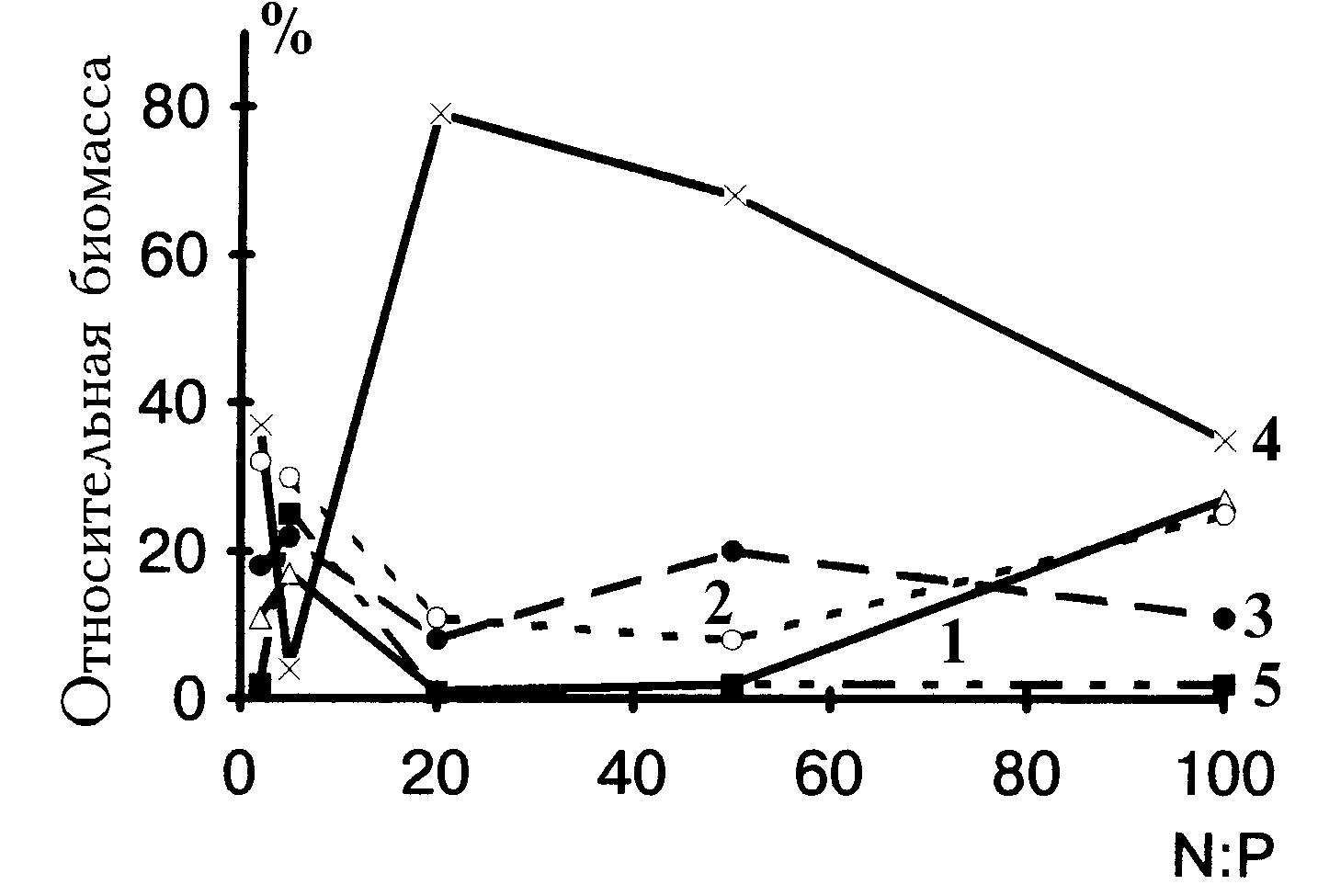
 С естественным прудовым фитопланктоном in vitro проводились два опыта в различные сезоны (Левич и др., 1992). Прудовое сообщество содержало более сотни видов, принадлежащих к нескольким крупным таксонам: Chlorophyta (Chlorococcales, Valvocales, Desmidiales), Bacillariophyta, Cyano-phyta
С естественным прудовым фитопланктоном in vitro проводились два опыта в различные сезоны (Левич и др., 1992). Прудовое сообщество содержало более сотни видов, принадлежащих к нескольким крупным таксонам: Chlorophyta (Chlorococcales, Valvocales, Desmidiales), Bacillariophyta, Cyano-phyta
Исследования in situ вели (Levich, Bulgakov, 1992; Левич и др. 1996 а, б) в течение нескольких лет на семи экспериментальных рыбоводных прудах. В опытные и контрольные пруды с начала апреля вносили минеральные азот и фосфор в различных пропорциях:
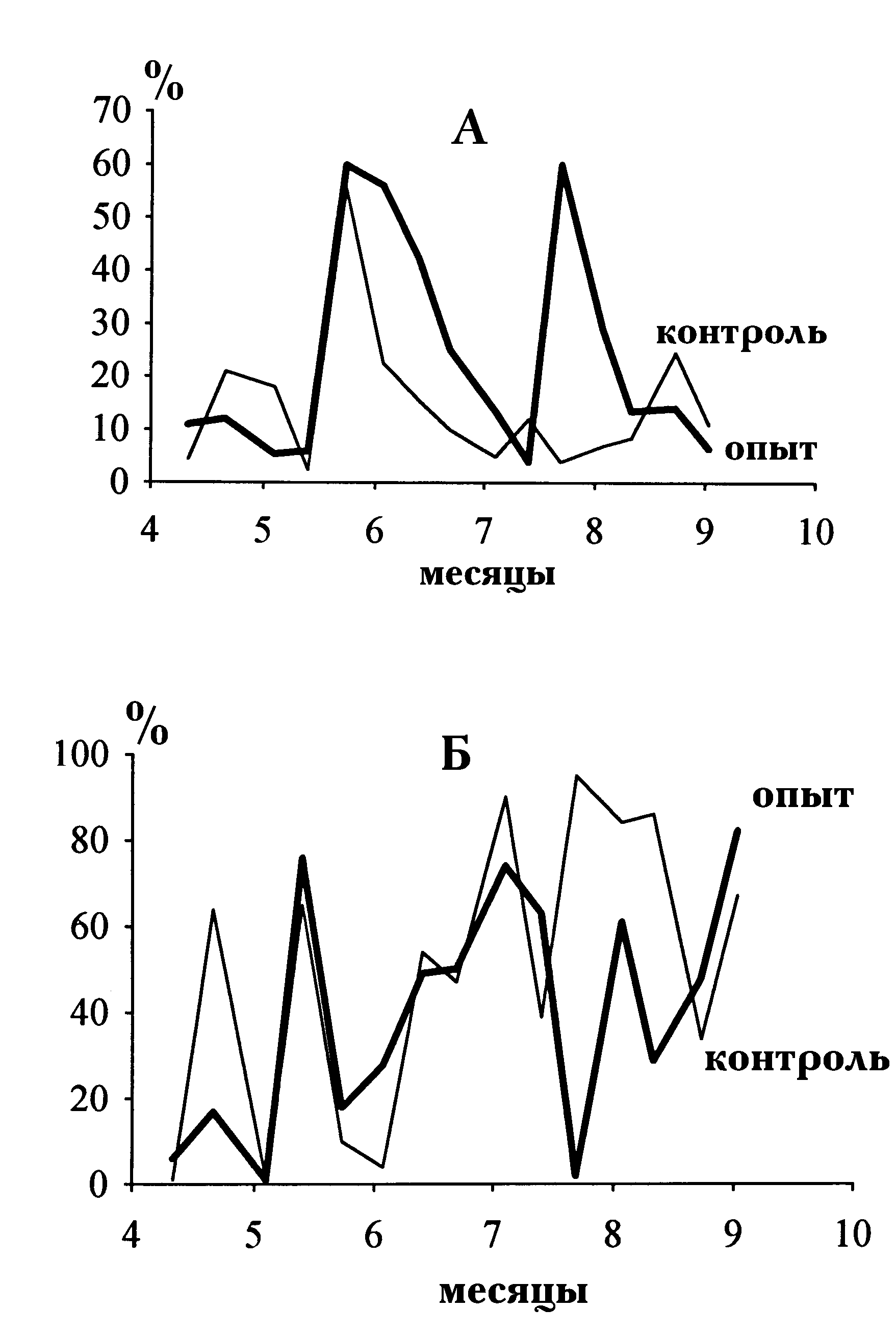
N/P, равное 4 в контрольных прудах, и N/P от 25 до 50 — в опытных. Кроме того, привлекались данные о фитопланктоне и гидрохимических характеристиках тех же прудов в другие сезоны (всего около 20 “годопрудов”). Исследовали биомассы популяций фитопланктона, контролировали гидрохимические показатели прудов. Высокие отношения N/P приводили к увеличению доли хлорококковых водорослей и снижению доли синезелёных (в составе которых доминировали неазотфиксирующие виды). Рис. 11 иллюстрирует типичную динамику относительных биомасс для одного из прудов в одном из сезонов. В таблице 3 приведены усредненные значения альгологических и гидрохимических характеристик прудов, демонстрирующие влияние элементов предложенной рациональной экологизированной системы удобрения на компоненты прудовой экосистемы. Среди этих элементов главенствующую роль играет повышение отношения количеств азота и фосфора во вносимых питательных веществах, а среди наблюдавшихся эффектов — повышение доли зеленых или крупных клеток в прудовом ценозе. Отсутствие значительной статистики опытных результатов в каждом из сезонов испытаний в определенной степени компенсируется воспроизводимостью эффектов в нескольких сезонах.
Таблица 3
Гидрохимические и гидробиологические параметры опытных и контрольных прудов за 1987-1989 гг.
|
Параметр |
1987 |
1988 |
1989 |
|||
|
опыт |
контроль |
опыт |
контроль |
опыт |
контроль |
|
|
Концентрация суммарного минерального азота в воде, мг/л |
||||||
|
средняя за апрель-май |
1.27 |
0.48 |
1.61 |
0.61 |
1.01 |
1.20 |
|
средняя за июнь-июль |
2.17 |
0.52 |
1.23 |
0.84 |
1.35 |
0.75 |
|
средняя за август-сентябрь |
2.74 |
0.89 |
1.30 |
1.18 |
3.50 |
1.73 |
|
среднесезонная |
1.94 |
0.57 |
1.40 |
0.86 |
1.81 |
1.16 |
|
Концентрация минерального фосфора в воде, мг/л |
||||||
|
средняя за апрель-май |
0.18 |
0.09 |
0.13 |
0.11 |
0.05 |
0.08 |
|
средняя за июнь-июль |
0.11 |
0.07 |
0.43 |
0.29 |
0.21 |
0.14 |
|
средняя за август-сентябрь |
0.10 |
0.10 |
0.37 |
0.24 |
0.09 |
0.08 |
|
среднесезонная |
0.13 |
0.08 |
0.30 |
0.21 |
0.13 |
0.10 |
|
Отношение концентраций азота и фосфора в воде |
||||||
|
средняя за апрель-май |
7 |
5 |
12 |
6 |
20 |
15 |
|
средняя за июнь-июль |
20 |
7 |
3 |
3 |
6 |
5 |
|
средняя за август-сентябрь |
27 |
9 |
4 |
5 |
39 |
22 |
|
среднесезонная |
15 |
7 |
5 |
4 |
14 |
12 |
|
Биомасса хлорококковых водорослей, мг/л |
||||||
|
средняя за апрель-май |
3.47 |
3.87 |
0.58 |
4.09 |
0.90 |
1.17 |
|
средняя за июнь-июль |
3.08 |
2.47 |
2.71 |
3.92 |
4.26 |
2.26 |
|
средняя за август-сентябрь |
5.13 |
5.01 |
11.18 |
9.39 |
4.92 |
3.13 |
|
среднесезонная |
3.65 |
3.32 |
3.83 |
5.60 |
3.32 |
2.13 |
|
Биомасса зеленых водорослей, мг/л |
||||||
|
средняя за апрель-май |
3.69 |
4.39 |
47.49 |
4.98 |
1.74 |
3.25 |
|
средняя за июнь-июль |
3.95 |
3.03 |
3.40 |
4.26 |
5.77 |
3.85 |
|
средняя за август-сентябрь |
8.24 |
8.02 |
14.53 |
9.66 |
6.34 |
4.09 |
|
среднесезонная |
4.98 |
4.48 |
24.15 |
6.14 |
4.58 |
3.71 |
|
Биомасса цианобактерий, мг/л |
||||||
|
средняя за апрель-май |
0.83 |
6.41 |
0 |
0 |
3.73 |
3.61 |
|
средняя за июнь-июль |
1.28 |
3.64 |
1.98 |
2.08 |
11.37 |
18.60 |
|
средняя за август-сентябрь |
5.64 |
4.08 |
2.43 |
3.35 |
22.15 |
17.34 |
|
среднесезонная |
2.30 |
4.16 |
1.27 |
1.60 |
11.7 |
13.27 |
|
Биомасса диатомовых водорослей, мг/л |
||||||
|
средняя за апрель-май |
4.48 |
10.00 |
0.91 |
11.68 |
3.38 |
1.22 |
|
средняя за июнь-июль |
2.36 |
3.86 |
0.76 |
1.34 |
1.79 |
1.30 |
|
средняя за август-сентябрь |
2.89 |
8.25 |
8.16 |
2.10 |
4.37 |
2.71 |
|
среднесезонная |
2.81 |
5.88 |
2.56 |
5.82 |
3.01 |
1.65 |
|
Биомасса эвгленовых водорослей, мг/л |
||||||
|
средняя за апрель-май |
6.71 |
8.78 |
5.61 |
3.32 |
0.12 |
0.14 |
|
средняя за июнь-июль |
4.25 |
7.22 |
1.07 |
0.83 |
1.46 |
0.51 |
|
средняя за август-сентябрь |
8.12 |
2.84 |
4.27 |
3.64 |
4.93 |
1.55 |
|
среднесезонная |
5.59 |
6.36 |
3.69 |
2.68 |
1.94 |
0.66 |
В дискуссиях с коллегами по поводу возможностей регулирования состава сообществ часто сквозит сомнение в том, что результаты конкуренции за ресурсы действительно определяются отношением содержаний ресурсов в среде, а не их абсолютными концентрациями. Поэтому сформулирую еще раз свой основной тезис: абсолютные дозы ресурсов безусловно определяют суммарную биомассу сообщества. Отношения количеств ресурсов определяют долю в общей биомассе, которая приходится на ту или другую (таксономическую, размерную) группу организмов, т.е. отношения ресурсов влияют на относительные биомассы.
Многочисленные лабораторные опыты с проточным культивированием музейных штаммов и природного фитопланктона обнаруживают существование оптимальных видоспецифичных отношений для концентраций азота, фосфора и кремния (Tilman, 1982; Sommer, 1983; Kilham, 1986; Suttle, Harrisson 1988; Grover, 1989; Suttle et al., 1991; De Pauw, Naessens-Foucquaert, 1991; Holm, Armstrong, 1991; Egge, Heimdal, 1994; Grover, 1997).
Широко известна серия многолетних экспериментов на малых озёрах Швеции (Schindler, 1977; Findley, Kasian, 1987; 1994), где изменение отношения
N/P также приводило к смене типа цветения водоёма. Зависимость высокого обилия синезелёных от низких отношений N/P (или высокого обилия зелёных при высоких значениях N/P) была замечена в озёрах США (McQueen, Lean, 1987; Stockner, Shortreed, 1988), Юго-Восточной Азии (Cho et al., 1990), Южной Африки (Haarhoff et al., 1992), Франции (Michard et al., 1996). Обратно пропорциональная зависимость между численностью синезелёных водорослей и величиной N/P показана путём статистического корреляционного анализа для многих озёрных экосистем (Varis, 1991).
Концепция направленного регулирования типов “цветения” эвтрофных водоемов
Модельные и эмпирические данные о роли отношений
N/P для видовой и размерной структур фитопланктонных сообществ позволили сформулировать концепцию направленного регулирования типов цветения водоёмов с помощью биогенного манипулирования (Левич, 1995a; Levich, 1996). Регулирование с помощью отношения N/P предоставляет при деэвтрофировании возможность парадоксального управления. А именно, обычный путь деэвтрофирования состоит в сниженнии фосфорной нагрузки (линия АВ на рис. 4). Но если для доминирования цианобактерий действительно существенно низкое отношение N/P, а не абсолютные дозы биогенных веществ, то добиться перехода от цветения водоема синезелёными к цветению, например, протококковыми можно и добавками в водоёмы азотных соединений (линия АС на рис. 4), что технически и экономически гораздо доступнее, чем изъятие фосфора или его ограничение в стоках. Возникающая при повышенных дозах питательных веществ дополнительная первичная продукция (теперь уже в виде биомассы хлорококковых) легко может быть утилизирована и превращена во вторичную продукцию консументами (например, специально индуцируемыми в водоём фитопланктоядными рыбами), поскольку зелёные водоросли в отличие от многих таксонов синезелёных не являются трофическим тупиком и, как правило, не токсичны. Переход из области однофакторного лимитирования (точка А на рис. 4) в область двухфакторного (точка С на рис. 4) к тому же гарантирует, что биогенные вещества в среде не будут накапливаться ввиду их полного сбалансированного потребления.Регуляция состава фитопланктонного сообщества может оказаться полезной и в случае оптимизации (по видовому или биохимическому составу, по размерной структуре) кормовых фитопланктонных сообществ при выращивании беспозвоночных, растительноядных рыб и других форм аквакультуры (Левич и др., 1996a; Левич и др., 1996б). Увеличение доли синезелёных с помощью снижения отношения
N/P в среде может оказаться полезным в целях обеззараживания сточных вод при их использовании для полива или в целях улучшения биологических свойств поливных вод.Подбор видов планктонных водорослей для сбалансированной утилизации многобиогенной нагрузки на водоем
Расположение областей лимитирования в пространстве ресурсов полностью зависит от значений потребностей
Формула видовой структуры и адекватность вариационной модели фитопланктонных сообществ
Вариационная задача имеет решение
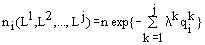 ,
,
задающее зависимость численностей ni видов сообщества от запасов лимитирующих рост ресурсов Lk и называемое формулой видовой структуры. Полная численность сообщества n
зависит от абсолютных концентраций Lk. Коэффициенты l k зависят от отношений Lr/Ls. Величины n и l k отыскиваются из системы алгебраических нелинейных уравнений, включающей балансовые ограничения вариационной задачи для лимитирующих ресурсов и соотношение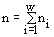 .
.
В прикладной экологии формула видовой структуры позволяет рассчитывать:
• численности ni и n, если заданы запасы ресурсов Lk и потребности ![]() ;
;
• парциальные потребления лимитирующих ресурсов среды популяциями сообщества ![]() , если заданы суммарные потребления ресурсов
, если заданы суммарные потребления ресурсов ![]() всем сообществом
всем сообществом ![]() ;
;
• парциальные ![]() и полные
и полные ![]() реальные потребления нелимитирующих ресурсов;
реальные потребления нелимитирующих ресурсов;
• потребности видов ![]() , если известны численности популяций ni и потребления Lk (в m сериях опытов для m ресурсов);
, если известны численности популяций ni и потребления Lk (в m сериях опытов для m ресурсов);
• формы ранговых распределений численности в сообществах, вводя зависимость этих распределений от распределений потребностей видов в лимитирующих ресурсах;
• параметрические индексы видового разнообразия сообществ, отражающие степень доминирования многочисленных видов, или резкость, выровненность, равномерность, однородность (и т.п.) убывания численностей в ранговых распределениях.
Таблица 4
Результаты предсказания с помощью общего набора квот относительной численности видов в сообществах (re — ранг вида в экспериментальном сообществе, rm — ранг вида в сообществе по модели, rq — ранг квоты, S — ранговый коэффициент корреляции по Спирмену)
|
Номер опыта |
|||||||||||
|
1 |
2.1 |
2.2 |
2.3 |
4.2 |
|||||||
|
rq |
re |
rm |
re |
rm |
re |
rm |
re |
rm |
re |
rm |
|
|
Ankistrodesmus braunii |
4 |
3 |
3 |
||||||||
|
Ankistrodesmus acicularis |
9 |
4 |
6 |
7 |
7 |
7 |
7 |
7 |
7 |
4 |
4 |
|
Scenedesmus obliquus |
2 |
1 |
1 |
||||||||
|
Scotiella nivalis |
8 |
5 |
5 |
5 |
6 |
6 |
6 |
6 |
6 |
||
|
Chromochloris cinnoborina |
12 |
9 |
9 |
||||||||
|
Chlamydomonas humicola |
13 |
10 |
10 |
||||||||
|
Chlorella vulgaris |
5 |
7 |
4 |
3 |
3 |
4 |
3 |
3 |
3 |
3 |
3 |
|
Stichococcus mirabilis |
11 |
8 |
8 |
8 |
8 |
8 |
8 |
8 |
8 |
||
|
Scenedesmus bijugatus |
10 |
6 |
7 |
||||||||
|
Scenedesmus quadricauda |
3 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
1 |
1 |
|
Ankistrodesmus falcatus |
6 |
6 |
4 |
3 |
4 |
4 |
4 |
2 |
2 |
||
|
Anacystis nidulans |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
||||
|
Pleurochloris magna |
7 |
4 |
5 |
5 |
5 |
5 |
5 |
||||
|
S |
0.915 |
0.929 |
0.976 |
1.000 |
1.000 |
||||||
В опыте с поликультурой из 10-ти видов зелёных микроводорослей (опыт №1) на стационарной фазе роста распределение численности клеток описывалось формулой видовой структуры ![]() с коэффициентом множественной корреляции 0,97 (Левич и др. 1986). Эта же серия опытов наряду с ещё шестнадцатью сериями экспериментов была проанализирована с помощью ранговых критериев адекватности выборок и с помощью калибровки клеточных квот (Замолодчиков и др., 1993). Серии опытов отличались друг от друга наборами таксонов (от четырех до десяти видов в поликультуре) и начальными концентрациями азота и фосфора в среде. При лимитировании роста единственным ресурсом место вида в ранжированном по убыванию ряду численностей на стационарной фазе согласно формуле видовой структуры полностью определяется рангом клеточной квоты вида по лимитирующему ресурсу (наибольшая численность соответствует наименьшей клеточной квоте). Была сделана попытка подобрать в пределах экспериментальной ошибки единственный набор рангов измеренных в опыте клеточных квот так, чтобы воспроизвести экспериментальные ранги численностей уже во всех сериях, где рост был ограничен единственным ресурсом. Это удалось сделать с ранговым коэффициентом корреляции Спирмена в пределах от 0.92 до 1.00 (табл. 4). Для серий с неоднофакторным лимитированием процедура проверки адекватности формулы видовой структуры состояла в следующем. По численностям видов в паре серий рассчитывали клеточные квоты по азоту и фосфору, а для всех остальных серий с этими откалиброванными квотами по формуле видовой структуры рассчитывали численности видов и сравнивали их с экспериментальными значениями. Сравнение показало, что рассчитанные значения численностей совпадают с измеренными в опыте в пределах экспериментальных ошибок.
с коэффициентом множественной корреляции 0,97 (Левич и др. 1986). Эта же серия опытов наряду с ещё шестнадцатью сериями экспериментов была проанализирована с помощью ранговых критериев адекватности выборок и с помощью калибровки клеточных квот (Замолодчиков и др., 1993). Серии опытов отличались друг от друга наборами таксонов (от четырех до десяти видов в поликультуре) и начальными концентрациями азота и фосфора в среде. При лимитировании роста единственным ресурсом место вида в ранжированном по убыванию ряду численностей на стационарной фазе согласно формуле видовой структуры полностью определяется рангом клеточной квоты вида по лимитирующему ресурсу (наибольшая численность соответствует наименьшей клеточной квоте). Была сделана попытка подобрать в пределах экспериментальной ошибки единственный набор рангов измеренных в опыте клеточных квот так, чтобы воспроизвести экспериментальные ранги численностей уже во всех сериях, где рост был ограничен единственным ресурсом. Это удалось сделать с ранговым коэффициентом корреляции Спирмена в пределах от 0.92 до 1.00 (табл. 4). Для серий с неоднофакторным лимитированием процедура проверки адекватности формулы видовой структуры состояла в следующем. По численностям видов в паре серий рассчитывали клеточные квоты по азоту и фосфору, а для всех остальных серий с этими откалиброванными квотами по формуле видовой структуры рассчитывали численности видов и сравнивали их с экспериментальными значениями. Сравнение показало, что рассчитанные значения численностей совпадают с измеренными в опыте в пределах экспериментальных ошибок.
Вывод
ыОптимальные для данного вида отношения ресурсов в среде (то есть те, при которых относительное обилие вида больше, чем при всех других отношениях) близки к отношениям потребностей этого вида в соответствующих ресурсах.
Управляющая роль отношений ресурсов в среде может быть востребована в мероприятиях по деэвтрофированию водоемов и в биотехнологиях по созданию предпочтительной кормовой базы для выращивания аквакультуры.
благодарности
Я глубоко благодарен В.Н.Максимову и Н.Г.Булгакову, без сотрудничества с которыми эта работа не была бы выполнена. Я признателен за поддержку и дружбу коллективам кафедр гидробиологии и общей экологии Биологического факультета Московского государственного университета. Представленная работа косвенно содержит труд многих моих соавторов, коллег, учеников и помощников. Это — В.Л.Алексеев, В.И.Артюхова, А.А.Боголюбова, Л.С.Болдырева, Д.Г.Дерягин, Д.Г.Замолодчиков, И.Е.Зелепукина, А.Б.Лебедь, Е.Г.Личман, Е.Д.Любимова, Н.З.Мазитова, Р.С.Никонова, В.А.Никулин, Н
.В.Новикова, В.Н.Носов, А.Л.Постнов, Н.В.Ревкова, Л.А.Риттих, С.Ю.Рыбакова, Т.В.Саломатина, Н.А.Смирнов, А.Ф.Сокольский, А.В.Соловьев, Л.Н.Тесленко, Е.А.Титова, Ю.Н.Торхов, И.Ю.Фенева, А.А.Худоян, А.И.Шаповалов, Н.А.Шидловская, В.В.Элиас. Всем им я искренне благодарен. Также я рад возможности выразить теплую признательность коллегам, которые принимали участие в обсуждении работы на разных ее этапах — В.А.Абакумову, В.В.Алексееву, Г.П.Гаврилову, А.М.Гилярову, Д.О.Логофету, Ю.Э.Романовскому, Г.Ю.Ризниченко, В.Д.Федорову, Ю.А.Шрейдеру и зарубежным экспертам В. Ламперту, М. Мунавару, В. Смиту, У. Зоммеру, М. Страшкрабе и Ц. Рейнольдсу.список работ, опубликованных по теме диссертации
Левич А.П.
Структура экологических сообществ // Биологические науки. 1977. №10. С. 63-74.Федоров В.Д., Кондрик Е.К., Левич А.П. Ранговое распределение численности фитопланктона Белого моря // Доклады АН СССР. 1977. T. 236. №1. С. 264-267.
Левич А.П. Экстремальный принцип в теории систем и видовая структура сообществ // Проблемы экологического мониторинга и моделирование в экологии. Труды ИПГ. Л.: Гидрометеоиздат.1978. Том 1. С. 164-183.
Левич А.П. Ранговые распределения численности фитопланктона Белого моря // Биологические науки. 1979. №4. С. 14-26.
Левич А.П. Структура экологических сообществ. М.: Изд-во Моск. ун-та. 1980. 181 с.
Федоров В.Д., Левич А.П. Откуда берутся индексы разнообразия? // Человек и биосфера. М.: Изд-во Моск. ун-та. 1980. Вып.4. С. 164-184.
Левич А.П. Теория множеств, язык теории категорий и их применение в теоретической биологии. Учебное пособие. М.: Изд-во Моск. ун-та. 1982. 190 с.
Левич А.П. Много ли видов могут сосуществовать на одном субстрате? // Второй Всесоюзный съезд паразитоценологов. Тез. докладов. Киев. Наукова Думка. 1983. С. 183-184.
Левич А.П., Носов В.Н., Смирнов Н.А. Теоретико-категорная модель структуры экологического сообщества с учетом внешних факторов // Человек и биосфера. М.: Изд-во Моск. ун-та. 1983. Вып.8. С. 134-139.
Левич А.П., Федоров В.Д., Марташвили Г.Ш., Любимова Е.Д., Смирнов Н.А., Семин В.А. Изучение подходов к совместному культивированию зеленых планктонных водорослей // Труды Тбилисского университета. 1983. С. 209-217.
Левич А.П., Ревкова Н.В., Любимова Е.Д., Мазитова Н.З. О существовании лабораторных альгоценозов // Научные доклады высшей школы. Биологические науки. 1985. №1. С. 62-69.
Ревкова Н.В., Булгаков Н.Г., Левич А.П. Рост четырех видов зеленых планктонных водорослей в моно- и поликультурах и попытка расчета видовой структуры. Рукопись депонирована в ВИНИТИ. 1985. 10 c. №7259—В.
Левич А.П. Поиск уравнений динамики и собственное время систем // Математическое моделирование в проблемах рационального природопользования. Тезисы докладов 10 школы-семинара. Ростов-на-Дону. 1986. С. 109-110.
Левич А.П., Гаврилов Г.П., Столяров А.Ю. Адекватность альтернативных моделей в экологии сообществ // Экологический прогноз. М.: Изд-во Моск. ун-та. 1986. С. 190-196.
Левич А.П., Любимова Е.Д., Марташвили Г.Ш. Видовая структура и потребление субстратно-энергетических факторов в лабороторных альгоценозах // Экологический прогноз. М.: Изд-во Моск. ун-та. 1986. С. 69-103.
Левич А.П., Ревкова Н.В., Булгаков Н.Г. Процесс “потребление-рост” в культурах микроводорослей и потребности клеток в компонентах минерального питания // Экологический прогноз. М.: Изд-во Моск. ун-та. 1986. С. 132-139.
Левич А.П., Лебедь А.Б. Потребности биологических видов в компонентах питания и потребление экологическим сообществом факторов среды // Проблемы экологического мониторинга и моделирования экосистем. 1987. Том 10. С. 268-283.
Levich A.P.
What are the Possible Theoretical Principles in the Biology of Communities? // Lectures in Theoretical Biology. Tallinn: Valgus. 1988. Pp. 121-128.Артюхова В.И., Быкова Н.И., Горюнова С.В., Левич А.П.
, Марташвили Г.Ш. Кинетика роста, потребление и потребности в азоте и фосфоре четырех видов планктонных водорослей // Вестник МГУ. Сер. 16. Биология. 1988. №1. С. 47-52.Левич А.П. Возможные пути отыскания уравнений динамики в экологии сообществ // Журнал общей биологии. 1988. Т.49. №2. С. 245-254.
Левич А.П., Булгаков Н.Г., Саломатина Т.В. Гидробиологическое состояние рыбоводного пруда в ответ на учащенное внесение минеральных удобрений // Тр. ГосНИОРХ. Л. 1988. Том 280. С. 143-149.
Левич А.П. Потребности фитопланктона в ресурсах среды и пути управления структурой альгоценозов // Журнал общей биологии. 1989. Т.50. №3. С.315-328.
Левич А.П., Титова Е.А., Личман Е.Г., Васин А.А. Модельный анализ управления структурой альгоценозов // Математические модели рационального природопользования. Ростов-на-Дону. 1989. С.5-15.
Левич А.П., Артюхова В.И. Измерение потребностей фитопланктона в субстратных факторах среды // Известия АН СССР. 1991. №1. С.114-123.
Levich A.P., Bulgakov N.G.
Regulation of species and size composition in phytoplankton communities in situ by N/P ratio // Russian Journal of Aguatic Ecology. 1992. №2. Pp. 149-159.Замолодчиков Д.Г., Булгаков Н.Г., Гурский А.Г.
, Левич А.П., Чесноков С.В. К методике применения детерминационного анализа для обработки экологических данных // Научные доклады высшей школы. Биологические науки. 1992. №7. С.116-133.Замолодчиков Д.Г., Левич А.П. Подбор видов планктонных микроводорослей для полной утилизации многобиогенной нагрузки на водоем // Вестник Моск. ун-та. Сер.16 Биология. 1992. №3. С.40-44.
Левич А.П., Личман Е.Г. Модельное изучение возможностей направленного изменения структуры фитопланктонного сообщества // Журнал общей биологии. 1992. Т.53. №5. С.689-703.
Левич А.П., Худоян А.А., Булгаков Н.Г., Артюхова В.И. О возможности управления видовой и размерной структурами сообщества в экспериментах с природным фитопланктоном in vitro. 1 // Биологический журнал Армении. 1991. Т. 44. №3. С. 186-193.
Левич А.П., Худоян А.А., Булгаков Н.Г., Артюхова В.И. О возможности управления видовой и размерной структурами сообщества в экспериментах с природным фитопланктоном in vitro. 2 // Научные доклады высшей школы. Биологические науки. 1992. №7. С.17-31.
Булгаков Н.Г., Левич А.П., Никонова Р.С., Саломатина Т.В. О связи между экологическими параметрами и продукционными показателями выростного рыбоводного пруда // Вестник Моск. ун-та. Сер.16 Биология. 1992. №2. С.57-62.
Levich A.P.
Toward a dynamic theory // Lectures in Theoretical Biology. V.2. Tallinn: Estonian Academy of Science. 1993. Pp. 33-50.Замолодчиков Д.Г., Левич А.П., Рыбакова С.Ю.
Исследование адекватности теоретико-категорной модели фитопланктонных сообществ // Проблемы экологического мониторинга и моделирования экосистем. Т.15. Л.: Гидрометеоиздат. 1993. С.234-246.Левич А.П. Теоретическая биология: поиск уравнений обобщенного движения // Известия РАН. Сер. биол. 1993. №2. С.320-323.
Левич А.П., Алексеев В.Л., Рыбакова С.Ю. Оптимизация структуры экологических сообществ: модельный анализ // Биофизика. 1993. Т.38. Вып.5. С.877-885.
Левич А.П., Булгаков Н.Г. О возможности регулирования видовой структуры лабораторного альгоценоза // Известия РАН. Сер. биол. 1993. №4. C.569-578.
Левич А.П., Замолодчиков Д.Г., Алексеев В.Л. Правило лимитирующего звена для многовидовых экологических сообществ // Журнал общей биологии. 1993. Т.54. №3. С.282-297.
Левич А.П., Алексеев В.Л., Никулин В.А. Математические аспекты вариационного моделирования в экологии сообществ // Математическое моделирование. 1994.Т.6. №5. С.55-71.
Levich A.P.
Time as variability of natural systems: ways of quantitative description of changes and creation of changes by substantial flows // On the Way to Understanding the Time Phenomenon: the Constructions of Time in Natural Science. Part 1. Interdisciplinary Time Studies. Singapore, New Jersey, London, Hong Kong: World Scientific. 1995. Pp.149-192. (Левич А.П. Время как изменчивость естественных систем: способы количественного описания изменений и порождение изменений субстанциональными потоками // Конструкции времени в естествознании: на пути к пониманию феномена времени. Часть 1. Междисциплинарное исследование. М: Изд-во Моск. ун-та. 1996. С.233-288.)Левич А.П. Критерии самоорганизации и управляющие параметры в экологии сообществ // Труды международной конференции “Критерии самоорганизации в физических, химических и биологических системах”. Суздаль. 1995. С.108-113.
Левич А.П. Экологические подходы к регулированю типов цветения эвтрофных водоемов // Доклады Академии наук. 1995. Т.341. №1. С.130-133.
Левич А.П., Булгаков Н.Г. Биогенные элементы в среде и фитопланктон: отношение азота к фосфору как самостоятельный регулирующий фактор // Успехи современной биологии. 1995. Т.15. Вып.1. С.13-23.
Левич А.П., Булгаков Н.Г., Никонова Р.С. Рациональное удобрение рыбоводных прудов с разновидовой посадкой // Известия РАН. Сер. биол. 1996. №1. С. 121-124.
Levich A.P.
The role of nitrogen-phosphorus ratio in selecting for dominance of phytoplankton by cyanobacteria or green algae and its application to reservoir management // J. of Aquatic Ecosystem Health. 1996. V.5. Pp.1-7.Левич А.П.
Феноменология, применение и происхождение ранговых распределений в биоценозах и экологии как источник идей для техноценозов и экономики // Математическое описание ценозов и закономерности технетики. Абакан: Центр системных исследований. 1996. С.93-105.Левич А.П., Булгаков Н.Г., Замолодчиков Д.Г. Оптимизация структуры кормовых фитопланктонных сообществ. Под редакцией проф. В.H.Максимова. М.: Товарищество научных изданий КМК. 1996. 136 с.
Левич А.П., Соловьев А.В. Категорно-функторное моделирование естественных систем // Анализ систем на пороге XXI века. М.: Интеллект. 1997. С.66-78.
Levich A.P., Solov'yov A.V.
Category-functor modelling of natural systems // Cybernetics and Systems. 1999. V.30. №6. Pp. 571-585.Alexeyev V.L., Levich A.P.
A search for maximum species abundances in ecological communities under conditional diversity optimization // Bull. of Mathemat. Biology. 1997. V.59. №4. Pp.649-677.Bulgakov N.G.
, Levich A.P. The nitrogeN/Phosphorus ratio as a factor regulating phytoplankton community structure //Archiv für Hydrobiologie. 1999. V.146. №1. Pp. 3-22.Левич А.П.
, Алексеев В.Л. Энтропийный экстремальный принцип в экологии сообществ: результаты и обсуждение // Биофизика. 1997. Т.42. Вып.2. С.534-541.Левич А.П., Булгаков Н.Г. Потребности планктонных водорослей в субстратных и энергетических ресурсах среды: концепция и измерения // Успехи современной биологии. 1997. Т.117. №1. С.107-121.
Левич А.П., Максимов В.H., Булгаков Н.Г. Теоретическая и экспериментальная экология фитопланктона: управление структурой и функциями сообществ. М.: Изд-во НИЛ. 1997. 192 с.
Левич А.П. Оптимизационное моделирование многокомпонентных открытых систем в условиях конкуренции за многочисленные ресурсы // Тезисы 5 Международной конференции “Математика, компьютер, образование”. М. 1998. С.118.
Levich A.P.
Variational modelling theorems and algocoenoses functioning principles // Ecological Modelling. 2000. V. 131. №2-3. Pp. 207-227.Левич А.П. Экстремальные принципы и вариационное моделирование для открытых сложных систем // Тезисы 2 Международной конференции “Актуальные проблемы современного естествознания”. Калуга. 2000 (в печати).