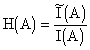 ,
, Энтропийный экстремальный принцип в экологии сообществ: результаты и обсуждение
Левич А.П., Алексеев В.Л.
Согласно предложенному энтропийному экстремальному принципу в реальности осуществляются состояния биологической системы, на которых достигается экстремум функционала, имеющего смысл меры структурированности системы. В частности, показано, что наличные ресурсы среды однозначно определяют состояние сообщества согласно полученной формуле видовой структуры. Сформулировано правило лимитирующего звена согласно которому для любой комбинации ресурсов в пространстве потребляемых ресурсов среды существуют области, в которых состояние сообщества полностью определяется только этой комбинацией. В частности, существуют области, где определяющим является любой фиксированный ресурсный фактор, любые два или несколько факторов сразу. Т.о. правило лимитирующего звена обобщает классические концепции лимитирования Либиха и Митчерлиха. Доказано, что с математической точки зрения рассматриваемый экстремальный принцип эквивалентен принципу минимального потребления, требующему минимизации потребления ресурса среды при ограничении на степень структурированности системы. Рассмотрен вариант энтропийного экстремального принципа, позволяющий рассчитывать траектории системы, как функции от потребленных ресурсов.
При описании функционирования биологических систем в теоретической биологии широко используется понятие экстремальности: принимается гипотеза, в силу которой из всего возможного спектра состояний биологической системы осуществляется то, для которого экстремальна некоторая заранее выбранная характеристика системы [1]. Например, в экологии в качестве такой характеристики используется показатель экспансии размножения популяций — тотальная биомасса [2], продуктивность [3] или удельная скорость роста [4]. В работах одного из авторов [5,6] на основании теоретико-категорных методов сравнения математических структур, был выдвинут энтропийный экстремальный принцип, который использовался затем в серии работ. В настоящей работе мы хотели бы привести некоторые результаты, полученные в последнее время при использовании этого принципа.
Энтропийный экстремальный принцип
Коротко напомним суть энтропийного экстремального принципа. Экологическое сообщество моделируется с помощью множеств с разбиениями [5,6]. Элементы множеств — это особи, классы разбиения — популяции биологических видов. Переходы сообщества из одних состояний в другие описываются соответствиями между множествами с разбиениями. Обычно из постановки задачи ясно, какие допускаются соответствия. Например, если мы хотим описать смертность особей, то допускаются не всюду определенные соответствия между множествами с разбиениями; если мы описываем размножение особей, то рассматриваются многозначные (нефункциональные соответствия) и т.д. С каждым состоянием сообщества, т.е. с соответствующим множеством с разбиением
A, мы свяжем следующую величину, называемую энтропией состояния A:,
где
Приведем здесь лишь некоторые из возможных интерпретаций экстремального принципа.
1) Разбиение множества
2) Величина
3) Как показано одним из авторов [5], при определенном выборе допустимых соответствий между множествами с разбиениями функционалы
H(A) совпадают с так называемыми императивными индексами разнообразия, используемыми в экологии. С этой точки зрения экстремальный принцип означает, что осуществляется состояние сообщества с максимальным разнообразием (см. также [7]).4) Энтропию системы можно рассматривать как количество информации, связанной со структурой системы [8], поэтому введенный экстремальный принцип можно интерпретировать и в информационных терминах.
В дальнейшем изложении будут обоснованы еще две интерпретации энтропийного принципа: принцип минимального потребления и энтропийная параметризация собственного времени системы.
Рассмотрим теперь сообщество, состоящее из организмов одного трофического уровня без возрастной структуры (например, из клеток фитопланктона), принадлежащих
w видам и потребляющих m взаимонезаменимых ресурсов. Пусть в рассматриваемом сообществе допустимы деление клеток и их гибель, но недопустимы поглощение одних организмов другими и интродукция особей извне. Таким требованиям удовлетворяют инъективные, сюръективные, не всюду определенные и не функциональные соответствия. Энтропийный функционал для такого сообщества приобретает вид [6]: ![]()
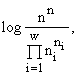 (1)
(1)
Вариационная задача включает ресурсные ограничения, которые имеют вид
![]() (2)
(2)
Здесь
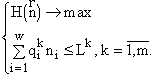 (3)
(3)
Решение задачи (3) методом Лагранжа приводит к формуле видовой структуры [5,10]
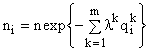 , (4)
, (4)
связывающей численности видов в сообществе с ресурсами
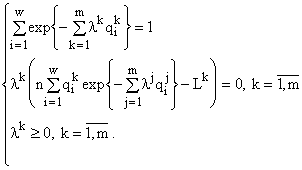 (5)
(5)
В работах [11,12] адекватность формулы видовой структуры установлена в опытах с лабораторными альгоценозами из 4, 8, и 10 видов, культивировавшихся на двух десятках сред с различными комбинациями начальных концентраций азота и фосфора в питательных средах.
Некоторые следствия энтропийного экстремального принципа
1) Вектор ресурсов
2) Из формулы (4) мы видим, что
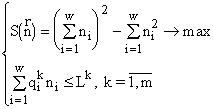
с функционалом
3) Так как вектор ресурсов
Пусть
m = 1. Тогда из первого уравнения (5) мы находим, что множительРазберем подробно случай
m = 2, который наиболее часто встречается в эксперименте и доказательство для которого содержит все идеи общей ситуации, будучи свободно от техническмх трудностей последней (теорема о максимуме видовых обилий).Введем обозначения
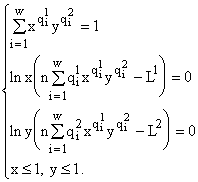 (6)
(6)
Из первого уравнения (6) мы можем найти
y как функцию от x. Примерный график y(x) представлен на рис.1. Из него видно, что для выполнения условий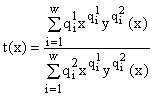 ,
,
где
y(x) находится из первого уравнения (6). Используя при вычислении dt/dx теорему о неявной функции, можно показать, что dt/dx > 0, т.е. t(x) строго монотонно возрастающая функция. Поэтому мы имеем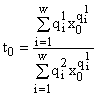
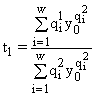 ,
,где
Таким образом, мы видим, что если вектор
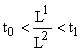 , то для вида i относительная численность
, то для вида i относительная численность ![]() ,
,
т.е. если
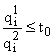 , то
, то 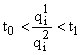 , то при
, то при 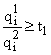 , то при t =
, то при t =Пусть теперь вектор
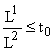 . Тогда в (6) надо положить x = x0, y = 1 и
. Тогда в (6) надо положить x = x0, y = 1 и 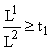 , то надо положить x = 1, y = y0 и
, то надо положить x = 1, y = y0 и В итоге мы получаем, что относительные численности зависят лишь от отношения
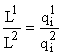 . Типичная картина поведения относительных численностей
. Типичная картина поведения относительных численностей Этот вывод остается справедливым и в общем случае: относительная численность заданного вида принимает наибольшее значение при отношениях в среде ресурсных факторов, равных отношениям потребностей в них данного вида.
Обзор многих экспериментальных фактов, согласующихся с этим выводом можно найти в работах [9,13,14,15,16].
4) Для задачи с неравенствами (3) доказано [10] правило лимитирующего звена, обобщающее принцип минимума Либиха и закон совокупного действия факторов Митчерлиха (подробности см., например, в [17]). Согласно этому правилу (теорема стратификации) пространство потребляемых ресурсных факторов среды распадается (стратифицируется) на
а) потребляются полностью;
б) являются аргументами экстремали (4) (т.е.
Таким образом, мы можем сказать, что состояние сообщества
Энтропийный экстремальный принцип и “теорема Гиббса”
Сделаем еще одно пояснение о смысле энтропийного экстремального принципа. Имеет место следующая теорема:
Пусть ресурсный фактор единственен и пусть
l — единственный корень уравнения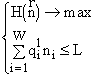
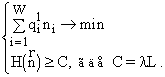
Эту теорему мы назвали “теоремой Гиббса”, т.к. в термодинамике аналогичное утверждение об эквивалентности задачи на максимум энтропии при фиксированном уровне энергии задаче на минимум энергии при фиксированном уровне энтропии доказал Гиббс [18].
Т.о. мы видим, что смысл энтропийного экстремального принципа достаточно прозрачен: требуется минимальность потребления ресурсного фактора при ограничении снизу на некоторую характеристику биосистемы, которую можно трактовать как ее сложность или разнообразие.
В случае
m ³ 2 возможно следующее обобщение “теоремы Гиббса”. Пусть мы имеем вектор 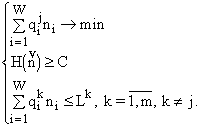 (7)
(7)
Тогда можно показать, что решение (4) задачи (3) является одновременно решением системы (7).
Верно и обратное утверждение: если
Экстремальный принцип, отраженный задачей (7), мы назовем принципом минимального потребления для любого из ресурсных факторов.
“Динамический” вариант энтропийного экстремального принципа
В естествознании экстремальные принципы традиционно применяются для получения уравнений движения. Достаточно упомянуть Лагранжев подход к механике, при котором уравнение траектории движения есть уравнение экстремали некоторого функционала (лагранжиана).
Как можно модифицировать энтропийный экстремальный принцип, чтобы получить какой-либо аналог траектории? Предлагается следующий принцип: из начального состояния
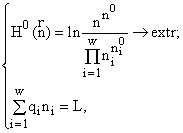 (8)
(8)
где
Легко видеть, что если
Анализ (8) показывает, что
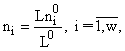 (9)
(9)
где
В многочисленных экспериментах с накопительными культурами фитопланктона установлено [21], что процесс роста численности клеточных популяций, обусловленный потреблением биогенного субстрата, закономеро складывается из трех стадий (рис.3):
Стадия A: Накапливание в клетках биогена без увеличения численности клеток.
Стадия B: Деление клеток за счет потребления вещества из среды.
Стадия C: Деление клеток за счет внутренних запасов питательного вещества.
Стадии могут быть редуцированными и перекрывающимися, но они имеют место как для всего сообщества в целом, так и для отдельных видов, входящих в его состав. Отметим, что стадии процесса рост-потребление лишь частично совпадают с фазами роста выявленными по кривым роста
n = n(t). А именно, стадия A есть заключительная часть лаг-фазы (на начальной части лаг-фазы нет ни роста культуры ни потребления вещества), а стадии B и C входят в фазу экспоненциального роста.Таким образом, мы видим, что на стадии B зависимость численности от потребленного субстрата линейна, т.е. аналогична (9). В этом случае
Мы надеемся посвятить одну из следующих работ более подробному анализу “динамического” энтропийного экстремального принципа.
Список литературы