Концепция и измерение потребностей фитопланктона
Концепция потребностей.
В настоящее время представления о потребностях клеток водорослей в компонентах питания, на наш взгляд, недостаточно разработаны. Сам термин “потребности” употребляется в различных смыслах. Отсутствует общепринятое научное определение данного понятия. Качественные представления о потребностях связаны с методом биологических испытаний (Брагинский, 1961): если добавка субстрата вызывает дополнительное продуцирование фитопланктона, признается, что клетки испытывают в этом субстрате потребность. За меру потребности клетки в субстратном факторе может быть принята (Общие основы изучения водных экосистем, 1979) константа полунасыщения (гиперболический закон Михаэлиса-Ментен), характеризующая зависимость скорости поглощения субстрата от его концентрации. Ряд авторов (см. Обзор: Максимова, 1977) связывают потребности клеток с концентрациями веществ в среде, обеспечивающими наилучшее развитие водорослей. Д.Тилман (Tilman, 1982) называет потребностью клеток ту концентрацию субстрата в среде, которая обеспечивает прирост численности или биомассы популяции, в точности компенсируемый смертностью особей. В качестве потребности фитопланктона в веществе можно также рассматривать альгологический аналог зоологического рациона — удельную скорость потребления питательного вещества. Эта скорость связана с первичной продукцией фитопланктона.Потребностью клетки в некотором веществе можно считать его содержание в единице биомассы или в одной клетке. Эту величину называют также внутриклеточной концентрацией вещества, или клеточной квотой (Droop, 1973). Роль потребности также играет “экономический коэффициент” ![]() — количество субстрата, необходимое для наращивания единицы клеточной биомассы.
— количество субстрата, необходимое для наращивания единицы клеточной биомассы.
В дальнейшем предлагается (Левич и др., 1986б) называть
потребностями величины
Таким образом, коэффициенты
Часто потребности видов в одних веществах оцениваются по известным потребностям в других с использованием стехиометрических отношений элементов в среде или в клетках; считается, что эти отношения постоянны. Например, для океанического фитопланктона приводится атомарное отношение элементов O:C:Si:N:P = 212:106:20:15:1 (Sverdrup et al.
, 1942). Однако в последнее время установлено, что благодаря способности фитопланктона создавать внутриклеточные запасы биогенных элементов, а также благодаря механизмам перестройки биохимического состава клеток при изменении условий их функционирования отношение веществ в клетках может варьировать. Так, С.Э.Йоргенсен (Jorgensen, 1979) для озерных сообществ микроводорослей приводит диапазон отношений азота к фосфору от 4.1 до 291. При помощи непосредственных авторадиографических методов обнаружено, что отношение внутриклеточных запасов углерода к запасам фосфора меняется у пресноводного фитопланктона от 3.3 до 500 (Gutelmacher, Petrova, 1982).К сожалению, в литературе отсутствуют систематические данные о количественных характеристиках внутриклеточных концентраций питательных веществ для отдельных видов фитопланктона. Столь же мало известна динамика клеточных квот.
В опытах по предварительному измерению потребностей квоты для десяти (Левич и др., 1986a) и для четырех (Артюхова и др., 1988) видов пресноводных зеленых водорослей рассчитывались как количества биогенного элемента, поглощенного из среды одной клеткой. Эти определения показали, что величины квот видоспецифичны; требуется количественное исследование зависимостей потребностей от факторов среды; на фоне роста культуры и изменения биогенных элементов в среде потребности крайне изменчивы и требуется выделение стадий процессов роста и потребления, на которых значения индексов более стабильны.
Стадии роста. В многочисленных лабораторных экспериментах, проведенных авторами (Левич и др., 1986б) с накопительными культурами фитопланктона, установлено, что процесс роста биомассы (численности) клеточных популяций, обусловленный потреблением биогенных элементов, закономерно складывается из трех стадий (рис. 3.1):
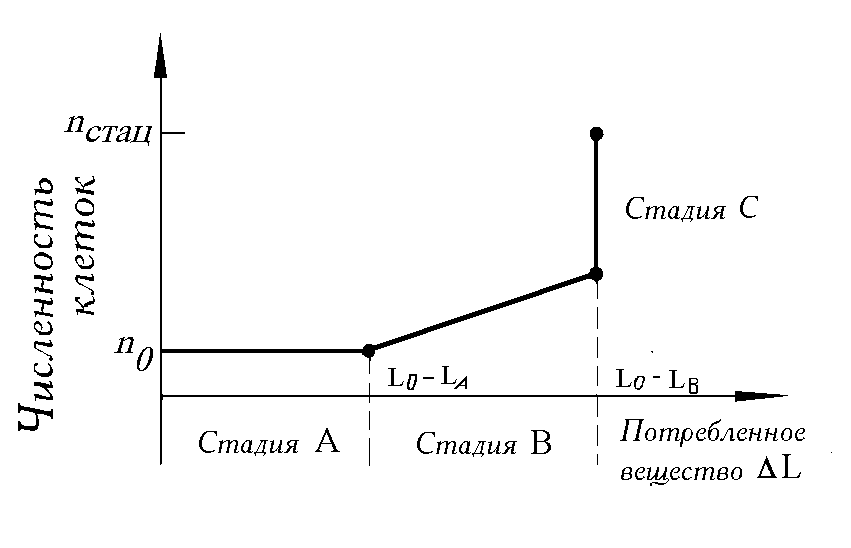
стадия A — поглощение веществ клетками и накопление биогенов без увеличения численности клеток;
стадия B — деление клеток при одновременном потреблении вещества из среды;
стадия C — деление клеток без потребления веществ из среды, т. е. за счет внутренних запасов питательного вещества.
Данные стадии, называемые составными частями процесса “потребление-рост”, могут быть перекрывающимися и редуцированными. Стадии могут лишь частично совпадать с фазами роста, выявленными по кривым роста. А именно, стадия A является составной частью лагфазы (на начальном этапе лагфазы нет ни потребления, ни роста), а стадии B и C входят в фазу экспоненциального роста.
При экспериментальном изучении роста популяции микроорганизмов стадия В наблюдается как при периодическом (накопительном), так и при проточном (непрерывном) культивировании, когда деление клеток происходит за счет веществ, поступающих в культиватор постоянно со свежей средой. Стадия С, как правило, составляет наибольшую по длительности фазу при накопительном культивировании. В природных водоемах рост популяции происходит в режимах, промежуточных между непрерывным и накопительным — приток биогенов может быть неоднократным в течение сезона, но завершается сезон, как правило, при отсутствии в среде веществ, лимитирующих рост.
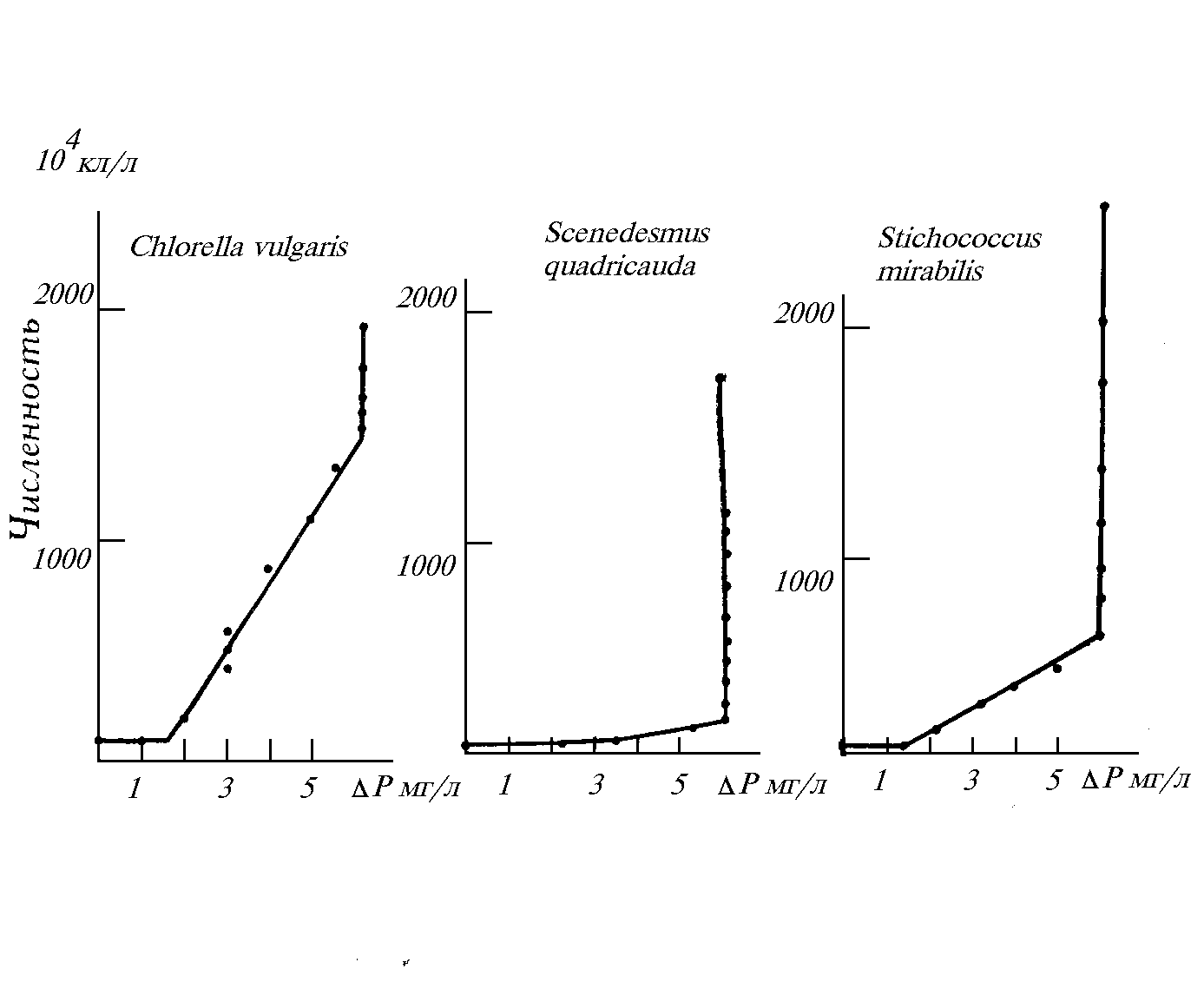
На рис. 3.2 в качестве иллюстраций приведены некоторые экспериментальные варианты идеализированной зависимости “потребление-рост” для ряда изученных видов в условиях лимитирования по фосфору.
Помимо относительной скорости роста клеточной популяции (vn = dn/ndt) и удельной скорости потребления биогенов (vL = dL/ndt), каждую стадию процесса “потребление — рост” удобно описывать следующими параметрами (Левич и др., 1986б):
Стадия A - начальная концентрация клеток n
- начальная концентрация биогена L
0;- содержание q
0 потребляемого вещества в каждой клетке в начале процесса;- содержание q
A потребляемого вещества в каждой клетке в конце стадии A;- концентрация вещества L
A, при которой начинается деление клеток (и заканчивается стадия A).Стадия B - количество вещества
lB, потребляемое из среды в расчете на одну вновь образованную клетку;Стадия C - количество питательного вещества q в каждой клетке;
- наименьшее содержание питательного вещества q
C в клетке, при котором прекращается деление клеток.Количество вещества
l, расходуемое любой клеткой при делении, и количество того же биогена q, содержащееся во вновь образованных клетках, связаны соотношением l=q+m, где m — количество биогена, выделяемое одной клеткой в среду при делении и в результате обмена веществ.Содержание вещества q в клетках в течение стадии B определяется не только веществом
lB, потребляемым из среды, но и долей биогена, доставшейся от материнской клетки:![]() .
.
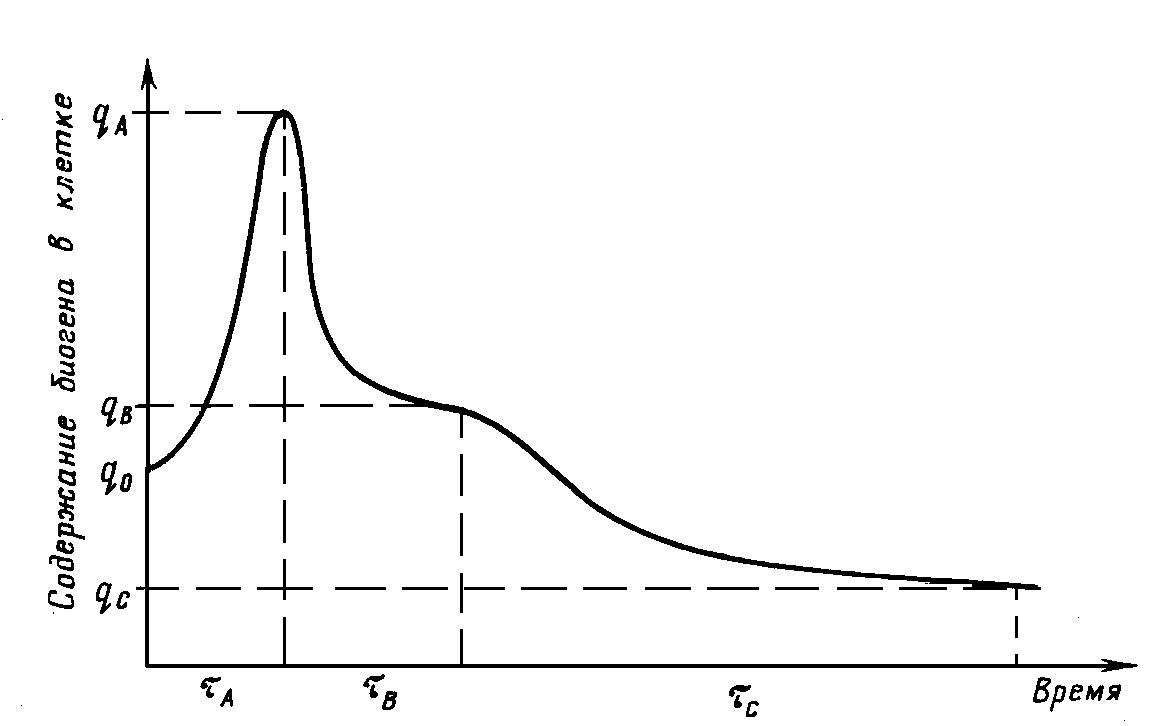
В течение стадии B не меняется количество вещества
lB, потребляемое из среды в расчете на одну вновь образованную клетку, и постоянно падает концентрация потребляемого биогена. На стадии C не изменяется пороговое значение биогена (как правило, близкое к нулю) и уменьшается содержание биогена в клетке. Динамика содержания вещества в клетке, фактически представляющая собой альтернативное рис. 3.1 описание существования стадий процесса “потребление — рост”, приведена на рис. 3.3.Эмпирическое соотношение
Приведем формулы расчета квот для каждой стадии процесса “потребление — рост”:

где nст — численность на стационарной стадии роста.
Анализ процесса “потребление-рост” показывает, что со сменой стадий происходит закономерное изменение самих потребностей: на стадии A вещество накапливается в клетках; при этом квота изменяется от начальной (qн) до максимальной (qмк). Во время стадии B потребление вещества из среды в расчете на одну клетку или единицу биомассы остается постоянным. Эта величина носит название квоты деления (qд). Как правило, qд меньше qмк. Это означает, что отсутствие клеточного деления на стадии A не связано с недостаточным для генерации роста внутриклеточным содержанием биогенного элемента. На стадии C квота постепенно уменьшается, и рост культуры останавливается по достижении минимальной квоты (qмн
).Методика измерения квот.
Содержание вещества в клетках можно было бы измерять прямыми химическими методами микроанализа. Однако существующие методики трудоемки, требуют большого количества клеточного материала, весьма чувствительны к погрешностям определения сухого веса вещества и обладают целым рядом других недостатков.Более показательным и сравнительно простым способом является определение квоты не прямым, а косвенным путем (Ревкова, 1987; Левич, Артюхова, 1991; Dauta et al., 1982). Реально измеряются убыль биогенного элемента из среды и прирост биомассы культуры за тот же отрезок времени. Квоты рассчитываются на основании закона о сохранении вещества: убыль вещества из среды принимается равной его потреблению клетками. Для определения начальной и минимальной квот необходимо довести культуру до стационарной фазы роста при двух различных исходных концентрациях ресурсов в среде:

где bн и bк — соответственно начальная и конечная биомасса.
Получается система двух уравнений с двумя неизвестными (qн и qмн), поскольку остальные величины известны из измерений. Подразумевается, что на обеих средах остановка роста произошла именно из-за нехватки вещества L. Если в двух рассматриваемых опытах лимитирующими будут разные элементы или, например, условия освещения, то конечная квота на стационарной фазе по данному элементу окажется не минимальной и может различаться в каждом из уравнений. Поэтому при определении квоты, например по фосфору, необходимо обеспечить отсутствие лимитирования по азоту, освещенности, температуре и т. д.
Обычно одну из двух исходных концентраций (пусть L
1) выбирают нулевой. Тогда![]() ;
; ![]() .
.
На стадии B, как отмечалось, отношение потребленного биогенного элемента к величине приросшей биомассы постоянно. Это отношение названо выше квотой деления и рассчитывается по измеренным в опыте на стадии B значениям
L и b:![]() .
.
Еще раз подчеркнем, что вычисленные таким образом квоты (особенно qд и qмн) не совпадают с точным содержанием вещества L в клетках, поскольку деление клеток сопровождается выделением веществ в среду. На стадии B, к примеру, содержание элемента в единице биомассы
![]()
можно сравнить с потребностью qд лишь при L >> b
н qн и при b >> bн. Таким образом, о величинах qд и qмн, называя их квотами, правильнее говорить как о количествах вещества, поглощенного из среды, а не содержащегося внутри клеток.Итак, клетки фитопланктона обладают специфическими потребностями в субстратных факторах среды. В таком случае не исключено, что существует также какой-либо показатель, характеризующий потребность водорослей в световой энергии.
Предложенный вариант оценки энергетических потребностей клеток основан на предположении о пропорциональности потребления световой радиации количеству ассимилированного углерода согласно соотношениям, задаваемым уравнением фотосинтеза. Б.Л.Гутельмахер (1986) исследовал статистическую связь между интенсивностью продуцирования органического вещества и объемом фотосинтезирующей клетки и пришел к выводу, что зависимость между суточной интенсивностью фотосинтеза (P/B-коэффициентом) и объемом клетки V, выраженным в миллиграммах углерода или биомассы на клетку, имеет вид:
![]() .
.
Коэффициент m зависит от систематического положения фитопланктона и водоема, в котором проходили исследования.
Таким образом, эффективность фотосинтетического процесса и эффективность утилизации световой энергии зависят от клеточного объема. Поэтому предлагается (Булгаков, 1990), установив потребление световой энергии пропорциональным углеродной продукции, называть потребностью в свете клетки величину q
E, прямо пропорциональную квадратному корню из клеточного объема (световая квота на единицу биомассы в этом случае обратно пропорциональна квадратному корню из клеточного объема).В качестве примера расчета потребностей фитопланктона в световой энергии рассмотрим результаты опытов с лабораторными поликультурами водорослей, где предположительно свет являлся лимитирующим фактором, поскольку в среду в течение эксперимента вносились биогенные элементы (Булгаков, 1990).
В опытах, где совместно выращивались
Scenedesmus quadricauda и Scenedesmus obliquus; Scenedesmus quadricauda и Ankistrodesmus falkatus, путем сравнения отношений квот по биогенам для каждого вида и отношения суммарных потреблений биогенов поликультурой было показано, что остановка роста культуры объясняется не только субстратным лимитированием, но и другими факторами (в данном случае — светом). Были построены балансовые уравнения как для азота и фосфора, так и для света: ![]() ; (1)
; (1)
![]() ; (2)
; (2)
![]() , (3)
, (3)
где q
N, qP и qE - квоты на единицу биомассы, измеренные в поликультурах соответственно для азота, фосфора и света; b1 и b2 — приросты биомассы для каждого вида за счет внешней среды за все время опыта.В правой части уравнений (1-3) стоит общее потребление фактора поликультурой. Находим из опыта
N и P, E вычисляем по формуле![]() ,
,
где vср — средний размер клеток поликультуры, равный
b/n;
b — общий прирост биомассы поликультуры за счет cреды;
n — соответствующий прирост численности;C — коэффициент из уравнения
![]() .
.
Тогда уравнение (3) примет вид:
![]() ,
,
или окончательно
![]() .
.
Из предположения, что квоты в моно- и поликультуре равны, следует вывод, что левая и правая части балансового уравнения должны совпадать в том случае, если фактор действительно был лимитирующим и квоты выбраны правильно.