Глава4
. Пути управления: эмпирические исследованияУправление структурой альгоценоза понимается как изменение относительных обилий таксономических или размерных групп фитопланктона при целенаправленном воздействии на сообщество управляющими факторами среды. Управление может оказаться полезным при оптимизации размерных, биохимических, токсикологических, трофических, продукционных характеристик фитоплактона в задачах регулирования типов цветения водоемов, обеспечения кормовых потребностей растительноядных рыб и беспозвоночных, а также в задачах осуществления биоэнергетических технологий.
Конечные прикладные цели предусматривают обычно регулирование состава альгоценоза в направлении доминирования клеток протококковых, диатомовых, эвгленовых и уменьшения доли сине-зеленых, а также увеличения доли крупных видов водорослей в общей биомассе.
Наряду с физическими и химическими методами подавления роста определенных таксонов фитопланктона возможен и чисто экологический подход, то есть управление жизнедеятельностью различных групп микроводорослей при помощи создания соответствующих условий, основанных на потребностях этих групп в ресурсных факторах среды. В частности, можно принципиально менять тип “цветения” путем внесения основных биогенных элементов в таких соотношениях, которые соответствовали бы клеточным квотам оптимизируемых групп фитопланктона по этим элементам (Левич,1989; Левич и др., 1992; Levich, Bulgakov, 1992).
В частности, одним из важнейших факторов, регулирующих структуру фитопланктона, является отношение незаменимых химических элементов — азота и фосфора — в питательной среде. Характер перестройки альгоценоза при смене этого отношения показан как для лабораторных опытов с чистыми культурами, так и для натурных и лабораторных экспериментов с природным фитопланктоном.
В разделе 3.5 описаны все возможные следствия изменения типа цветения водорослей в водоеме на примере их утилизации мирным зоопланктоном и растительноядными рыбами и на примере участия в процессах эвтрофирования.
4.1. Лабораторные альгоценозы
В ряде работ приведены результаты экспериментов, использующих методику непрерывного культивирования (см. 2.2). Данные опыты имели общую схему: путем различных добавок веществ в среду испытывали широкий набор отношений азота к фосфору, фосфора к кремнию и др., среди которых находились отношения, оптимальные для членов сообщества.
Поликультура из двух видов водорослей: диатомовой
Asterionella formosa и сине-зеленой Microcystis aeruginosa — выращивалась в режиме непрерывного культивирования (Holm, Armstrong, 1981). Диапазон атомных отношений кремния к фосфору, испытанных в опыте, варьировал от 2 до 200. Отношение биомасс видов с ростом отношения кремния к фосфору изменилось соответственно от 1:99 до 96:4. Оказалось, что A. formosa является видом, более требовательным к кремнию, но зато получающим преимущество перед конкурентом в условиях ограничений по фосфору. Примечательно, что сходные соотношения биомасс складывались при различных абсолютных концентрациях кремния и фосфора, но при близких отношениях этих элементов в среде. Например, водоросль M. aeruginosa могла преобладать при отношениях кремния к фосфору 25:2 (98% от общей биомассы) и 100:10 (97%), а A. formosa — при отношениях 300:2 (93%) и 100:0.5 (96%).При исследовании смешанной непрерывной культуры диатомей
A. formosa и Cyclotella meneghiniana (Tilman, 1977) применялся широкий диапазон отношений кремния к фосфо ру. При молярном отношении биогенных элементов < 6 преобладала A. formosa, а при отношении более 90 явно доминировала C. meneghiniana.Рост цианобактерии
Synechococcus nageli в поликультуре с зеленой Scenedesmus quadricauda изучался в условиях ограничения по фосфору, т. е. при высоких отношениях азота к фосфору (Suttle, Harrison, 1988). В такой ситуации S. nageli подавлял своего конкурента.Влияние температурных, световых и биогенных условий на исход конкуренции изучалось в опытах с культивированием двух морских водорослей
Skeletonema costatum и Phaeodactylum tricornutum. S.costatum вытесняла Ph. Tricornutum при низких значениях N:Si в среде и высоких — N:P (Pauw de, Naessens-Foucqaert, 1991). Оптимальное для S.costatum соотношение N:Si:P = 25:25:1.Различие в поведении видов при варьировании добавок лимитирующих веществ показано и на уровне монокультур. При сравнении урожайности двух видов зеленых водорослей
Scenedesmus quadricuada и Stigeoclonium tenue критическим для первого вида оказалось отношение азота к фосфору, равное 22, а для второго — 17 (Vries, Klapwijk, 1987). Критическим авторы называют такое отношение, превышение которого ведет к лимитированию роста фосфором.На основе опытов с монокультурами
Scenedesmus sp. Г.-Ю. Ри (Rhee, 1978) пришел к выводу, что для этой водоросли стимулирующим рост является отношение, равное 30; ниже и выше этого значения замечено ограничение роста культуры соответственно азотом и фосфором. Оптимальное отношение N:P, найденное для цианобактерии Anacystis nidulans, равно 30 (Сиренко, 1972).В серии экспериментов Г.-Ю.Ри и Дж.Готэм (Rhee, Gotham, 1980) обнаружили для монокультур водорослей видоспецифичность оптимального отношения азота к фосфору, содержащихся в питательной среде. Например, для диатомеи
Melosira binderana оно равно 7, для зеленой водоросли Scenedesmus obliquus — 30.К.Терри (Terry, 1985) измерил оптимальное отношение азота к фосфору для культуры водоросли
Pavlova lutheri, оказавшееся равным 43.На основе культивирования лабораторных штаммов микроводорослей в поликультурах были изучены (Левич, Булгаков, 1993) оптимальные отношения азота к фосфору в питательной среде и предпринята попытка управления видовым составом простейшего искусственного альгоценоза путем варьирования отношений исходных концентраций азота и фосфора.
Опыты проводились с накопительными культурами микроводорослей. Заданные в начале опыта дозы минеральных веществ по мере убыли из среды не возобновлялись. Численность клеток увеличивалась, пока не достигала стационарной фазы роста в результате субстратного лимитирования. О результатах конкуренции между видами судили по конечной (стационарной) численности каждого из них.
Поликультуры видов, участвующих в опыте, выращивались на среде Дота (Dauta, 1982а), содержащей несколько вариантов сочетаний азота (в виде KNO
3) и фосфора (в виде K2HPO4).В колбы с предварительно простерилизованной жидкой средой также стерильно инокулировались альгологически чистые музейные культуры водорослей. Полученные таким образом поликультуры выращивались в люминостате при постоянном люминесцентном освещении интенсивностью 7.3 Вт/м
2 за исключением опыта №1 (см. ниже). Продолжительность опытов определялась временем наступления стационарной фазы роста.В первом опыте (см. 2.3, опыт № 4) участвовали четыре вида протококковых водорослей
Scenedesmus quadricauda (Turp.) Brеb., Сhlorella vulgaris Beyer, Ankistrodesmus falcatus (Corda) Ralfs и Ankistrodesmus sp., поликультура из которых росла на двух средах со следующими начальными концентрациями биогенных элементов:среда 1 — 11 мг/л азота и 3 мг/л фосфора (N:P
3.5);среда 2 — 50 мг/л азота и 2.5 мг/л фосфора (N/Р
20).Искусственное сообщество второго опыта (см. 2.3, опыт №9) состояло только из двух видов протококковых
S. quadricauda и A. falcatus, но при большем наборе соотношений азота и фосфора:среда 1 — 4 мг/л азота и 3.1 мг/л фосфора (N/Р
1.3);среда 2 — 14 мг/л азота и 3.1 мг/л фосфора (N/Р
4.5);среда 3 — 34 мг/л азота и 0.6 мг/л фосфора (N/Р
57).В состав поликультуры в третьем опыте (№1) входили десять видов водорослей, из которых существенный вклад в биомассу поликультуры вносили восемь видов зеленых: порядок
Protococcales — Ch. vulgaris, Chromochloris cinnoborina Chodat., S. quadricauda, S. bijugatus (Turp.) Kutz., S. obliquus (Turp.) Kutz., Ankistrodesmus acicularis Korschic., A. braunii Brunnth.; порядок Ulothrichales — Stichococcus mirabilis Lagerh. Сообщество выращивалось на двух средах с одинаковым содержанием азота и фосфора — соответственно 195 и 5.2 мг/л, но при разных условиях освещенности — 0.15 и 4.4 Вт/м2.В течение каждого из опытов периодически под микроскопом определяли численность клеток. Параллельно проводили измерение содержания биогенных элементов в среде.
При обработке данных опытов рассчитывали начальные и конечные субстратные клеточные квоты для всех видов в соответствии с методикой определения потребностей клеток фитопланктона в азоте и фосфоре на разных стадиях роста (см. гл. 3). Вычисление световых потребностей и оценку потребляемой поликультурой световой энергии осуществляли на основе величин видоспецифических клеточных объемов (гл.1). Итоги роста видов в условиях искусственного альгоценоза оценивали по их парциальному вкладу в общую численность после остановки роста всех видов, входящих в поликультуру.
Для того чтобы понять, какая доля конечной численности каждого вида вырастала за счет минеральных ресурсов среды, во всех опытах оценивалась величина прироста численности за счет запасов азота и фосфора, депонированных в клетках к моменту начала опыта. Для этого предварительно вычисляли начальные клеточные квоты всех видов.
Продолжительность первого опыта составила 37 суток. Видимо, этого оказалось недостаточно для достижения стационарной фазы всеми видами. Впрочем, данное обстоятельство не заслонило тех эффектов, которые были обнаружены и которые в случае достижения остановки деления клеток, по-видимому, проявились бы более наглядно.

На обеих средах с учетом роста на запасах и субстратах среды в сообществе доминировал вид
S. quadricauda. Однако, если при отношении азота к фосфору, равному 3.5, это доминирование было не столь безусловным (44% от общей численности), то при увеличении отношения до 20 культура S. quadricau-da практически вытеснила три других вида из сообщества (рис. 4.1А). Относительная численность последних в сумме составила 17%. Абсолютная численность, например Ch. vulgaris, на среде с высоким отношением N:P в 6 раз меньше, чем на среде с низким отношением. При N:P 3.5 стационарной фазы роста достигли только популяции S. quadricauda и A. falcatus. При отношении, равном 20, наоборот, вид S. quadricauda был единственным, продолжавшим расти в момент окончания эксперимента. Поэтому доведение поликультуры до полной остановки деления скорей всего привело бы к усилению обнаруженной тенденции.Сравнение отношений конечных численностей видов, выросших только на азоте и фосфоре среды, показало, что при переходе от низкого отношения N:Р к высокому меняется доминирующий вид сообщества. Если в первой серии опытов при N:P
3.5 абсолютно доминирует Ch. vulgaris (100%), то при N:P 20 около 93% общей биомассы составляет биомасса S. quadricauda (рис. 4.1Б).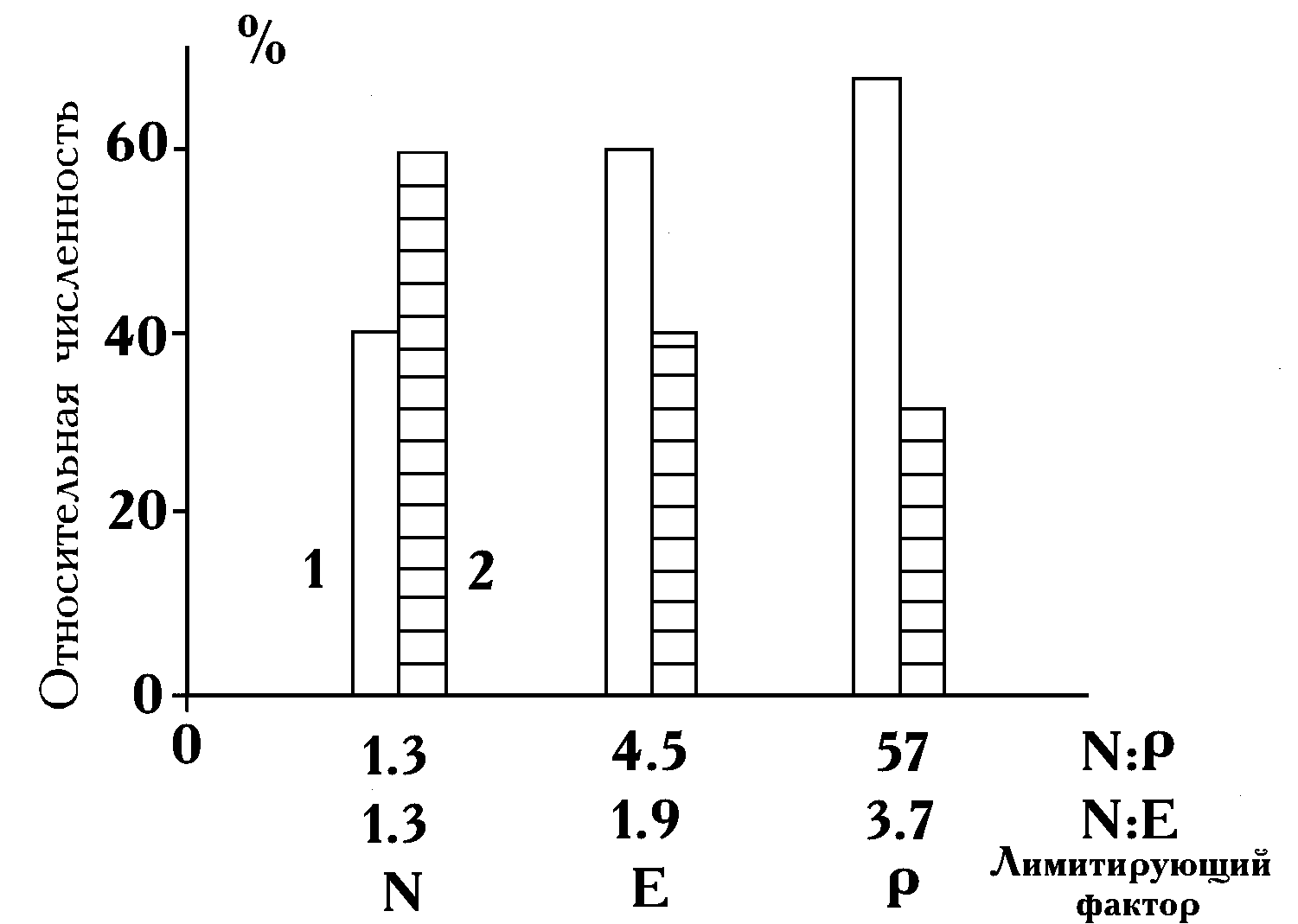
Результаты исследования роста двух видов в течение двух месяцев во втором опыте проиллюстрированы на рис. 4.2. При посеве инокулятов культур во всех средах поликультура состояла на 20% из клеток
S. quadricauda и на 80% — из A. falcatus.По мере увеличения отношения азота и фосфора меняется и соотношение конечных обилий двух видов.
S. quadricauda увеличивает свое видовое обилие и в лимитированных азотом средах (отношения 1.3 и 4.5), и в среде, ограниченной по фосфору (отношение 57). Изменение его относительной численности происходит при отношениях 1.3, 4.5 и 57 — соответственно от 40 до 60 и 68%. В этой серии рост культур на запасах был невелик по сравнению с общим приростом, поэтому соответствующие поправки не внесли существенных изменений в распределение биомасс видов на всех трех средах.В третьей серии опытов рост видов продолжался 68 суток, хотя при освещенности 0.15 Вт/м
2 некоторые виды достигли стационарной фазы раньше. Высокая исходная концентрация азота в среде привела к тому, что элемент к концу опыта не потреблялся поликультурой полностью и, следовательно, не мог быть лимитирующим. Поэтому в данном опыте рассматривалось влияние на видовую структуру альгоценоза отношения фосфора и световой энергии (Р:Е) как внешних факторов роста. Эти отношения были равны: в среде с высокой освещенностью — 0.35, в среде с недостатком света — 1 (методика вычисления величины E описана в гл. 3).Конечные видовые обилия всех популяций поликультуры, выросших без учета клеточных запасов, представлены в табл. 4.1. Видно, что при переходе освещенности от 4.4 Вт/м
2 к 0.15 Вт/м2 изменяется видовая структура сообщества. Относительная численность одних видов (A. acicularis, S. obliquus, S. bijugatus, S. quadricauda) уменьшается, других (Ch. vulgaris, Ch. cinnoborina, S. mirabilis) — увеличивается. Вклад A. braunii в общую численность практически не претерпевает изменений.Таблица
4.1Соотношение конечных численностей видов в восьмивидовой поликультуре при двух уровнях освещенности (%)
|
Вид |
Относительная численность, % при освещенности |
Отношение кво-ты по фосфору к |
||
|
4.4 Вт/м 2 |
0.15 Вт/м 2 |
квоте по свету |
||
|
Ankistrodesmus braunii |
22 ? |
23 |
0.5 |
|
|
A. acicularis |
21 |
16 |
0.4 |
|
|
Scenedesmus obliquus |
19 |
14 |
0.3 |
|
|
S. bijugatus |
5 |
0 |
0.5 |
|
|
S. quadricauda |
33 |
14 |
0.2 |
|
|
Chlorella vulgaris |
0 ? |
16 |
0.4 |
|
|
Chromochloris cinnoborina |
0 |
6 |
2 |
|
|
Stichococcus mirabilis |
0 |
11 |
0.8 |
|
|
Р:Е |
0.35 |
1 |
||
Примечание. Подчеркнуты отношения квот, наиболее близкие к единице. Выделены относительные численности тех видов, которые, исходя из отношения квот, должны иметь более высокую относительную численность в данной поликультуре. Около численностей видов, не отвечающих этому правилу, стоят знаки вопроса, соответствующее обсуждение приведено в тексте.
В табл. 4.1 также приведены отношения конечных клеточных квот по фосфору и по свету. Для обеих поликультур подчеркнуты относительные численности тех видов, которые должны были бы увеличивать свой вклад в суммарную численность на данной среде, исходя из гипотезы о близости отношения клеточных квот и отношения поглощенных сообществом ресурсов. Заметно, что виды с высоким отношением потребностей (0.8 — 2)
Ch. cinnoborina и S. mirabilis имеют и более высокий вклад в общую численность в условиях дефицита световой энергии. Остальные виды обладают отношением квот, близким к 0.35, но не все из них имеют более высокую относительную численность на среде, лимитированной по фосфору: видовые обилия А. braunii практически одинаковы на обеих средах, а Ch. vulgaris при освещенности 4.4 Вт/м2 вообще не росла.Величина световой квоты чувствительна к изменению объема клетки. Поэтому ошибка в определении линейных размеров могла привести к искажению значений квоты и отношения квот. По-видимому, реально клетки
A. braunii и Ch. vulgaris были мельче и соответственно отношение фосфорных и световых потребностей для этих видов было выше.S. quadricauda
получает преимущество перед другими членами сообщества при высоких отношениях азота к фосфору (от 20 в первой серии до 57 во второй). Преимущество S. quadricauda перед A. falcatus во второй серии связано с лимитированием по свету, и управление вызвано сменой отношения N:E.По-видимому, оптимальным для данного таксона микроводорослей можно признать отношение биогенных веществ в среде, равное их отношению непосредственно в клетке. Только в этой ситуации возможно сбалансированное потребление всех субстратов. В противном случае какой-либо из пищевых ресурсов будет потреблен раньше других, что приведет к преждевременной остановке роста.
В основе конкуренции и сосуществования лежат отличия оптимальных клеточных отношений двух субстратных факторов. Этот факт можно проиллюстрировать (Rhee, Gotham, 1980) следующей схемой. Допустим, для двух видов, составляющих поликультуру, оптимальные отношения азота к фосфору различны: у вида А оно равно 10, у вида В — 20. При каком-либо N:Р, находящимся в пределах от 10 до 20, система из двух видов находится в области сосуществования. Тогда добавление в среду фосфора будет менять отношение в сторону, благоприятную для вида А, снижая конкуренцию со стороны В, а азотная подкормка будет более благотворной для вида В, сопровождаясь постепенной элиминацией А. Одновременное добавление обоих ресурсов вызовет увеличение суммарной численности с неизменными видовыми обилиями.
Известно (см. гл. 3), что содержание биогенных веществ в клетках во время развития популяции непостоянно. Соотношение веществ, оптимальное для конечных численностей, очевидно, должно определяться конечными клеточными квотами или, в случае достижения культурой стационарной стадии роста, минимальными пороговыми потребностями клеток в компонентах минерального питания, достижение которых приводит к прекращению деления. При составлении поликультур, имеющих разные отношения минимальных квот по азоту и фосфору, преимущество в росте будут получать те из них, для которых отношение наиболее близко к отношению концентраций этих элементов в среде. Очевидно, что при ограничении роста альгоценоза недостатком освещенности преимущество в развитии должны получать виды, у которых отношение одной из субстратных квот к энергетической квоте близко к отношению потребляемых соответствующих внешних факторов.
Описанное выше поведение фитопланктонных сообществ можно предсказать с помощью математических моделей (см. подробнее гл. 2). При анализе некоторых из них (Левич, 1989; Левич, Личман, 1992; Левич и др., 1993) делаются три основных вывода: в качестве управляющих воздействий, формирующих относительные численности видов в альгоценозе, могут фигурировать отношения концентраций биогенных элементов в среде, а не сами абсолютные концентрации; в качестве видоспецифических характеристик клеток, определяющих отличия в поведении видов, выступают потребности видов в ресурсах среды; оптимальные для вида отношения ресурсов в среде равны характерным для вида отношениям соответствующих потребностей клеток в этих ресурсах.
Данные рассмотренных выше экспериментов, по-видимому, не противоречат этим выводам. Действительно, в первом опыте на первой среде рост поликультуры был ограничен азотом, а на второй среде — фосфором и ни на одной из сред, как показывают расчеты, не было недостатка света. Для доминировавшего (по приросту за счет веществ среды, а не внутриклеточных запасов) на первой среде (N:Р
3.5) вида Ch. vulgaris отношение потребности в азоте к потребности в фосфоре, по нашим данным и данным других авторов (Dauta, 1982б; Tezuka, 1985) для видов рода Chlorella, может находиться в пределах 7-10. А для доминировавшего на второй среде (N:Р 57) вида S. quadricauda это отношение в среднем заключено в диапазоне 20-30 (Левич, Артюхова, 1991; Muller, 1972; Dauta, 1982б).Во втором опыте рост видов ограничен на первой среде азотом, на второй — светом и на третьей — фосфором. В этой серии виды достигли стационарной стадии роста как в моно-, так и в поликультурах, в силу чего их конечные квоты по лимитирующему веществу должны быть равны минимальным и могли быть и были экспериментально измерены. Для вида
A. falcatus отношение потребностей в азоте и фосфоре, равное отношению соответствующих минимальных клеточных квот, оказалось равным 7, а для вида S. quadricauda — 25. Следовательно, при сравнении первой, лимитированной по азоту (N:Р 1.3), среды с третьей, лимитированной по фосфору (N:Р 57), нужно заключить, что анкистродесмус должен преобладать на первой среде, а сценедесмус — на третьей, что и наблюдалось в эксперименте (рис. 4.2) при сравнении первой лимитированной по азоту (N:E 1.25) среды со второй — лимитированной по свету (N:E 1.9). Сопоставим величину N:E с отношением потребностей видов в азоте и свете. Это отношение для А. falcatus равно 1.25 и для S. quadricauda — 1.5, что соответствует доминированию анкистродесмуса на первой среде, а сценедесмуса — на второй (рис. 4.2).В третьем опыте в условиях светового лимитирования лучше росли виды с высоким отношением потребностей в фосфоре и световой энергии. С ростом освещенности, наоборот, увеличивали свои парциальные численности виды с низким отношением Р:E в клетках.
4.2. Природный фитопланктон
in vitroКак показывают литературные данные, в природном фитопланктоне наиболее оптимальными для зеленых водорослей являются отношения азота к фосфору в среде, большие 20
25 (Smith, 1983). Напротив, сине-зеленые, как правило, занимают доминирующее положение в сообществе при отношениях N:P 510 (Schindler, 1977), а более высокие значения N:P часто ингибируют их рост. Показано, что стимулирующее или ингибирующее влияние соотношений биогенных элементов может носить и более узкую, видоспецифическую направленность.У.Соммер (Sommer, 1983), меняя в среде молярное отношение кремния к фосфору от 4 до 80, культивировал в хемостате природные многовидовые (более 30 видов) популяции фитопланктона из Боденского озера. При недостатке кремния доминировала водоросль
Mougeotia thylespora. При примерно сбалансированных отношениях самыми массовыми были виды Koliella spiculiformis, Synedra acus и Asterionella formosa. При недостатке фосфора подавляюще преобладала только Synedra acus. Эти результаты наблюдались вне зависимости от состава природного инокулята. Сравнение с видовыми обилиями фитопланктона в озере показало, что при отношениях биогенных элементов, близких к экспериментальному, в естественном водоеме исход конкуренции был аналогичным.С.Саттл и П.Харрисон (Suttle, Harrison, 1988) провели ряд экспериментов с природным пресноводным фитопланктоном в лабораторных условиях при отношениях азота к фосфору 5, 15 и 45. При отношении, равном 45, наблюдалось абсолютное доминирование цианобактерий рода
Synechococcus. При более низких отношениях наиболее многочисленными оказались диатомеи Nitzschia holsatica и Synedra radians и зеленая водоросль Scenedesmus sp.С.Килхэм (Kilham, 1986), анализируя рост фитопланктона из озера Мичиган в лаборатории при различных отношениях кремния к фосфору в среде, также заключает, что ответственным за видовую структуру альгоценоза оказывается не какой-то отдельный ресурс, а именно их парциальные количества. Он показал, что при лимитировании фосфором диатомеи вытесняют из сообщества представителей всех прочих отделов, но после снижения также и концентрации кремния доминирование переходит к зеленым. В средах с высоким отношением Si:P (313 и 71) преобладали диатомовые водоросли, в то время как при низких отношениях (4.6 и 0.9) — зеленые. Поскольку испытывались очень высокие отношения азота к фосфору (100), цианобактерии элиминировали в опыте.
К сходным выводам пришел Дж. Гровер (Grover, 1989) при полупроточном культивировании озерного сообщества водорослей. При отношении добавок в среду кремния и фосфора, равном 20, преобладали зеленые водоросли, а при Si:P
80 — диатомеи. Кроме того, при увеличении отношения до 160 в ценозе преимущественно развивались кремнежгутиковые водоросли.При инокулировании в полиэтиленовых мешках сообщества водорослей из озера Чит (Западная Вирджиния, США) с низким pH (доминирующий вид — синезеленая водоросль
Anabaena variabilis) увеличение значения pH и снижение N:P в среде приводило к увеличению биомассы фитопланктона без смены доминирования (Wilcox, De Costa, 1990). Показано, что значение N:P > 25 неблагоприятно для роста синезеленых.В озеро Спроут (Канада) вносили азотные и фосфорные удобрения в соотношении N:P = 50, после чего отбирали пробы и в лаборатории изучали кинетику потребления N и P водорослями двух размерных фракций (< 3 мкм и > 3 мкм). Соотношение количеств потребляемых N и P рассчитывали как соотношение максимальных скоростей поглощения этих элементов. Оказалось, что мелкая фракция (преимущественно
Synechococcus spp.) ограничена N, а крупная (преимущественно Rhizosolenia spp.) — P. Следовательно, для мелких видов оптимальным будет N:P > 50, а для крупных — < 50 (Suttle, Harrison, 1988).Нами были проведены эксперименты с природным фитопланктоном, где оценивалось влияние разных отношений минеральных форм азота и фосфора на видовую и размерную структуру природного альгоценоза рыбоводного пруда (Левич и др., 1992).
Таблица 4.2
Действующие факторы роста водорослей в опыте №1
|
№ аквариума |
Концентрация, мг/л |
Отношение азота |
||
|
азот |
фосфор |
к фосфору |
||
|
1 |
3.3 |
0.3 |
12 |
|
|
2 |
3.3 |
0.3 |
12 |
|
|
3 |
3.3 |
0.7 |
5 |
|
|
4 |
5.8 |
0.3 |
16 |
|
|
5 |
5.8 |
0.3 |
16 |
|
|
6 |
5.8 |
1.2 |
5 |
|
|
7 |
0.8 |
0.3 |
5 |
|
Опыты проводили в контролируемых условиях. Для этого воду из рыбоводного пруда помещали в шесть аквариумов объемом 20 л, в которые затем добавляли суперфосфат и аммиачную селитру в разных количественных сочетаниях. С учетом природного фона в двух аквариумах отношение N:P составило 5, в двух — 12, в двух — 16. Исходные биомассы каждого вида во всех аквариумах были одинаковы. Чтобы исключить эффект выедания зоопланктоном, опытную воду перед внесением добавок пропускали через ячеистую сеть с соответствующим размером ячей и оставляли на двое суток в темноте. В одном из аквариумов исходная концентрация биогенных элементов была равна фоновой. В течение опыта все аквариумы содержали на открытом воздухе. В табл. 4.2 указаны начальные концентрации азота и фосфора и их соотношения в каждом аквариуме с учетом природного фона. Для некоторых абсолютных концентраций и соотношений существуют повторности, по которым для всех фитопланктонных показателей проводились усреднения.

С помощью микроскопирования определяли численность и одновременно массы клеток фитопланктона (путем измерения индивидуальных размеров). Полученная биомасса служила основным функциональным показателем различных систематических групп. Наблюдения за динамикой роста водорослей вели в течение 14 суток, однако уже в середине опыта в аквариумах сильно возросла численность зоопланктона. Это произошло за счет того, что сквозь ячеи все же прошли покоящиеся яйца и мелкие формы зоопланктона. Поэтому примерно после 10-х суток опыта значения биомассы фитопланктона из-за интенсивного выедания нельзя признать функциями только питания и роста, так что в качестве конечных были использованы значения биомасс, усредненные по данным за 6-е и 10-е сутки. Следует отметить, что примерно к этому времени запасы фосфора и азота из среды были исчерпаны, и дальнейший рост был возможен только на внутриклеточных запасах и обороте органических форм этих элементов.
Анализ конечной биомассы вели как по крупным таксонам фитопланктона (порядки протококковых и вольвоксовых, отделы зеленых, диатомовых и сине-зеленых), так и на уровне родов и видов. Таксоны уровня рода и вида были поделены на доминирующие (их биомасса составляет не менее 20% от суммарной на шестые сутки опыта хотя бы в одном из аквариумов), непредставительные (биомассой — менее 1%) и субдоминирующие (все остальные).
Из рис. 4.3 видно, что стимулирующим воздействием на порядок протококковых обладает самое высокое отношение азота к фосфору, равное 16. Влияние этих условий заметно и на примере протококковых доминантов —
Scenedesmus acuminatus и рода Coelastrum (рис. 4.4). У других порядков и отделов увеличение отношения N:P ведет к замедлению роста. Самое заметное снижение скорости роста у цианобактерий наблюдалось при отношении N:P 16.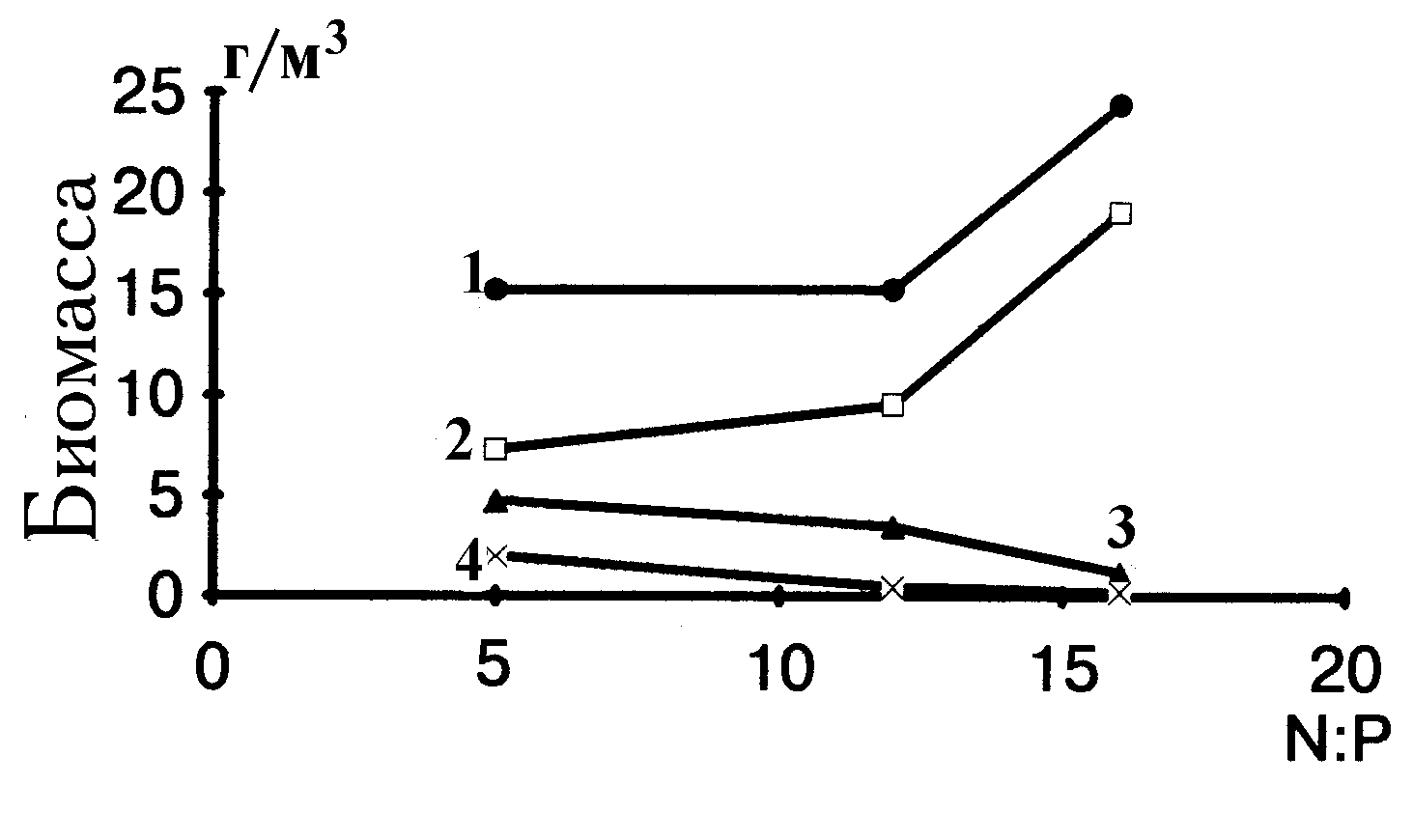
Следует подчеркнуть, что, хотя и можно выявить закономерности в изменении абсолютных биомасс крупных таксонов водорослей, действие отношения азота к фосфору является видоспецифичным. Так, доминирующие среди протококковых виды
Pediastrum duplex, S. quadricauda, S. acuminatus, род Coelastrum увеличивают свою биомассу при повышении отношения азота к фосфору (рис. 4.4), что и определяет увеличение биомассы протококковых в целом. В то же время протококковые водоросли родов Actinastrum, Ankistrodesmus, Dictiosphaerium существенно снижают биомассу при отношении, равном 16. Снижение биомассы сине-зеленых водорослей определяется представителями родов Phormidium и Merismopedia, а снижение биомассы диатомовых — представителями родов Nitzschia и Fragilaria (рис. 4.4). В то же время диатомовые рода Navicula наибольшей биомассы достигают именно при отношении N:P 16, хотя и не могут компенсировать снижение биомасс доминирующих родов этого отдела.Анализ данных размерной структуры включал в себя сравнение средних масс особи порядков и отделов (величина определяется путем деления суммарной биомассы таксона на суммарную его численность в одни и те же сутки опыта), а также сравнение относительных долей в биомассе определенных размерных классов водорослей. Для этого все найденные в пробах виды были разделены на 6 размерных групп, исходя из их объемов. После перевода объемных единиц в единицы массы получились следующие классы клеток: 1) менее 0.1 нг; 2) от 0.1 до 0.3 нг; 3) от 0.4 до 1 нг; 4) от 1.1 до 3.2 нг; 5) от 3.3 до 10 нг; 6) более 10 нг. Данное разбиение на размерные классы соответствует разбиению на равномерные отрезки на логарифмической шкале масс.
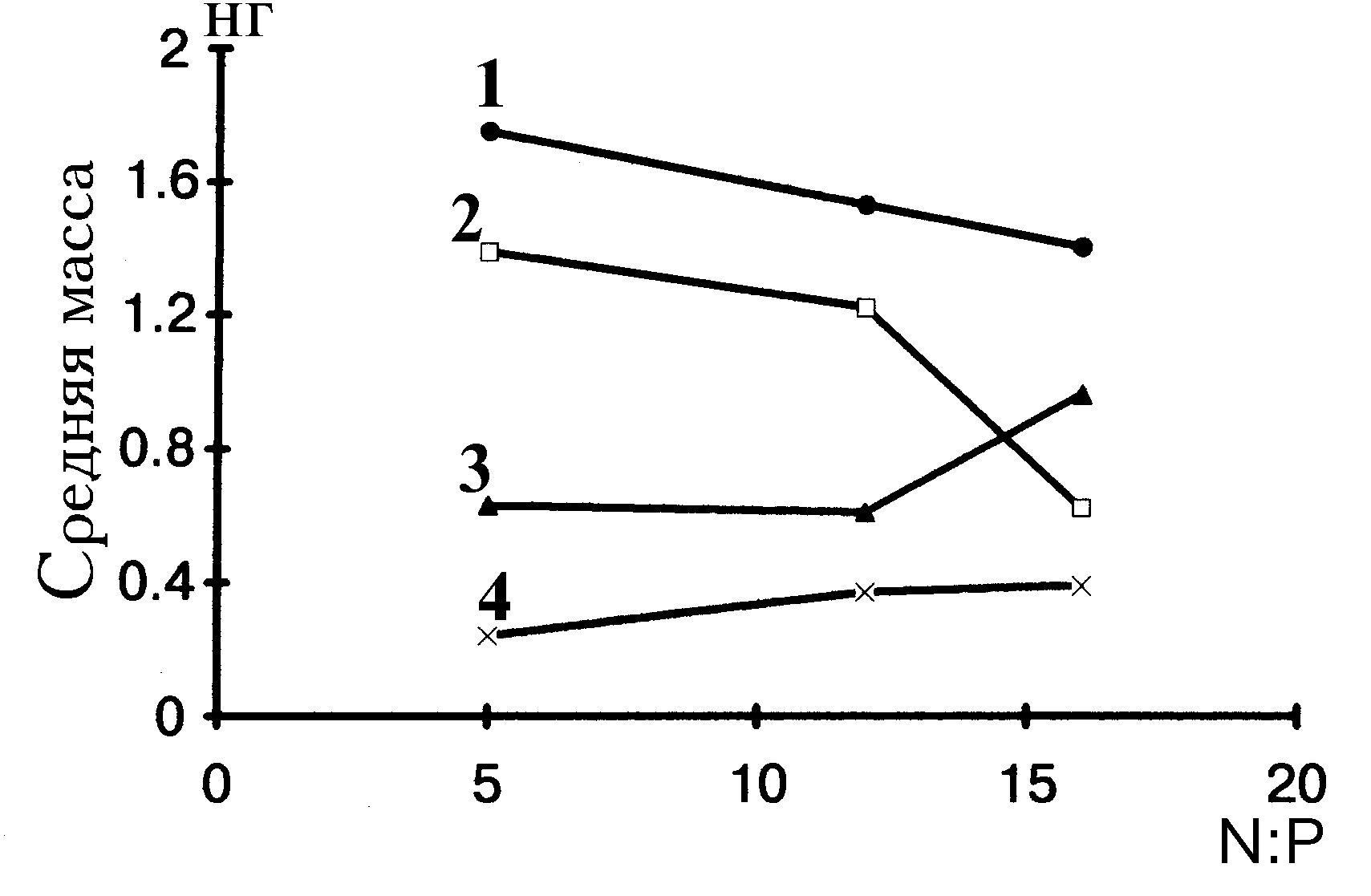
Результаты, приведенные на рис. 4.5, показывают, что повышение отношения азота к фосфору в среде закономерно уменьшает средние объемы клеток вольвоксовых и сине-зеленых и несколько увеличивает (при величине отношения, равной 16) средний объем клеток протококковых.
Следует пояснить, что в данном случае речь идет не об изменении абсолютных индивидуальных размеров клеток, а о смене доминирования в сообществе видов с разными размерами особей. Например, при повышении отношения азота к фосфору среди протококковых начинают преобладать виды с более массивными клетками.
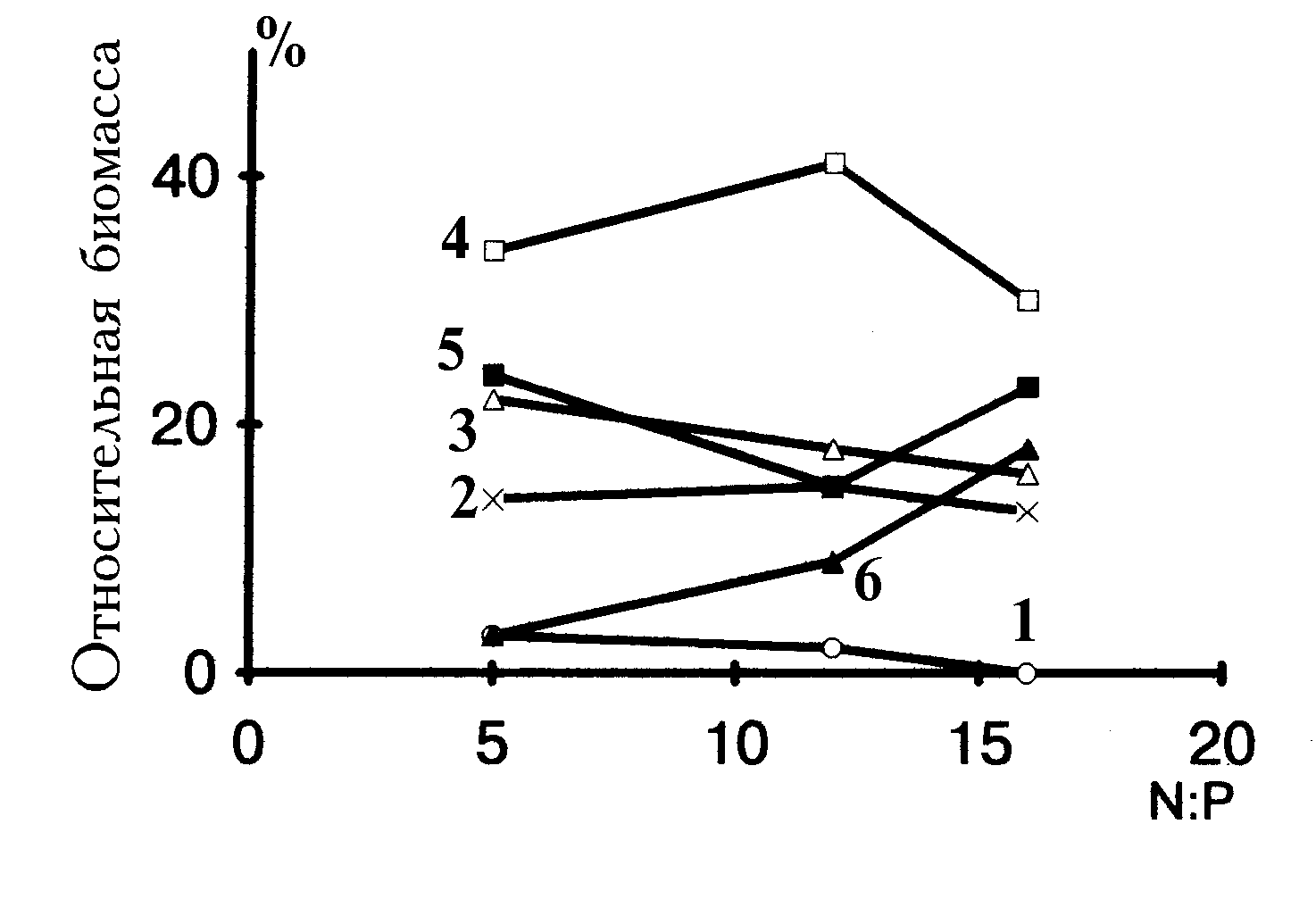
Влияние высоких отношений азота и фосфора на относительное обилие размерных классов проявляется только у видов с самыми крупными клетками (более 10 нг), которые повышают свою относительную биомассу (рис. 4.6).
В другом эксперименте нами была предпринята попытка проследить за реакциями таксонов фитопланктона на более широкий (по размаху и количеству) набор отношений N:P.
Схема второго опыта с более широким набором исходных отношений биогенных элементов (отношение N:P составляло 2, 5, 20, 50 и 100, табл. 4.3) не имела принципиальных отличий за исключением того, что постановку опытов осуществляли в двухлитровых колбах. Меры по изъятию зоопланктона оказались более эффективными и в течение всех восьми суток, пока длился опыт, водоросли не испытывали пресса выедания.
Таблица
4.3Действующие факторы роста водорослей в опыте №2
|
№ колбы |
Концентрация, мг/л |
Отношение азота |
||
|
азот |
фосфор |
к фосфору |
||
|
1 |
20 |
0.2 |
100 |
|
|
2 |
10 |
0.2 |
50 |
|
|
3 |
10 |
0.5 |
20 |
|
|
4 |
1 |
0.2 |
5 |
|
|
5 |
1 |
0.5 |
2 |
|
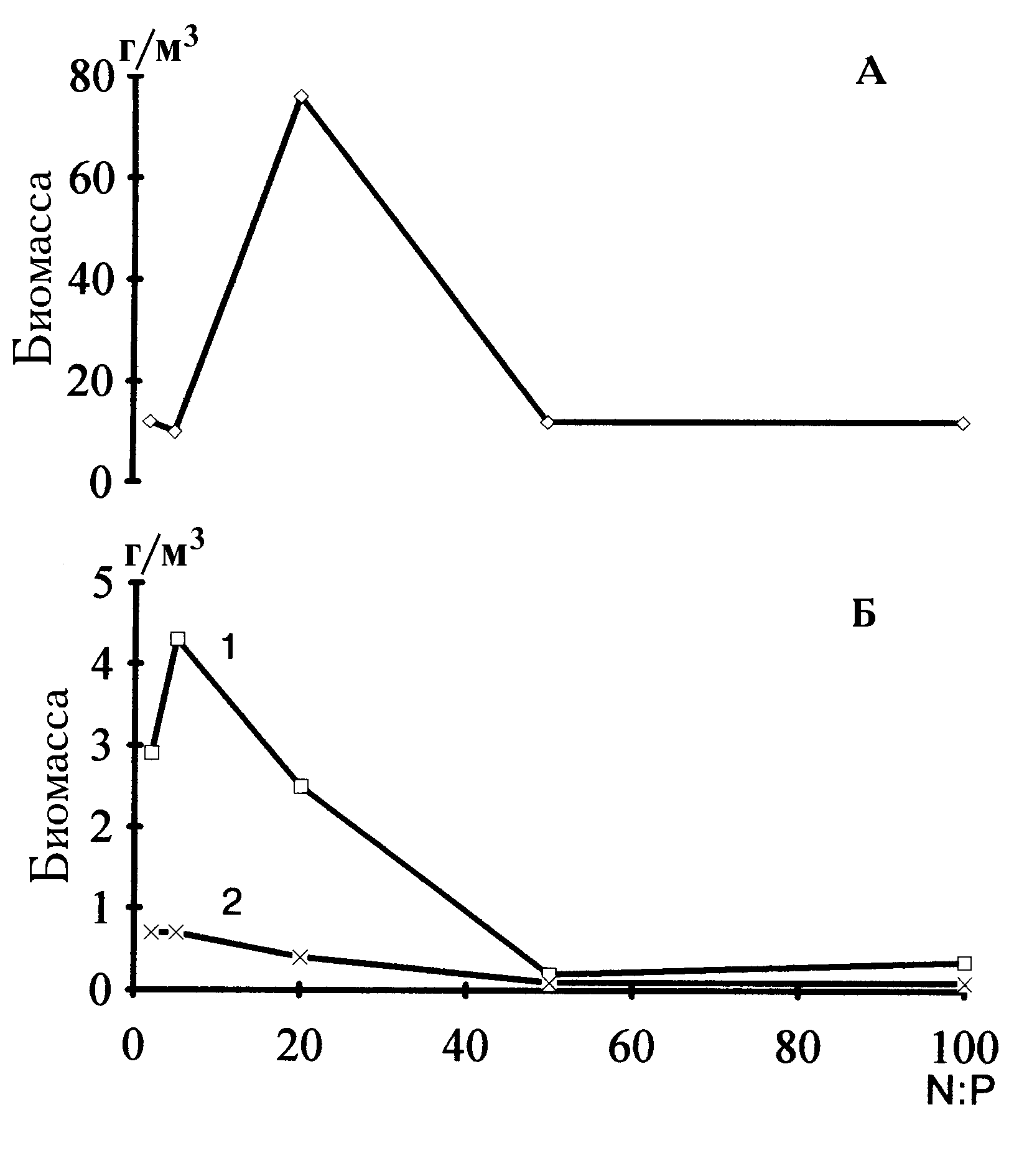
На рис. 4.7 приведены конечные биомассы для трех основных отделов фитопланктона в зависимости от начальных отношений биогенных элементов. Отношения N:P, большие 5, как видно, заметно преображают структуру альгоценоза в направлении абсолютного доминирования зеленых. Биомасса зеленых водорослей имеет один пик, соответствующий самому интенсивному росту. Этот пик приходится на отношение азота к фосфору, равное 20. У диатомей и цианобактерий максимальная биомасса достигается при низких отношениях (2
5). Увеличение азотных добавок влечет за собой угнетение развития.

Сравнение поведения доминирующих видов и родов из этих отделов показывает, что биомасса
S.quadricau- da практически полностью определяет поведение всего отдела зеленых (рис. 4.8). Другой представитель зеленых род Didymocystis имеет и второй пик биомассы при отношении 100. Таким образом, при более широких пределах варьирования отношения азота к фосфору заметна специфика и в откликах доминирующих видов зеленых. Для родов диатомей Stephanodiscus и Nitzschia оптимальными являются отношения в диапазоне от 5 до 20. Наконец, сине-зеленая водоросль Microcystis sp. наилучшим образом развивается при отношениях 25. Более высокие отношения выступают для нее в качестве ингибирующего фактора. В то же время сине-зеленая водоросль Anabaena sp. увеличивает биомассу при высоких отношениях.На рис. 4.9 показано, как реагируют на разные отношения биогенных элементов средние размеры особей основных отделов на восьмые сутки опыта. Самые крупные клетки зеленых были обнаружены в колбе с отношением 20. При переходе к более высоким отношениям средние размеры снижаются, но все же остаются большими, чем при отношениях 2 и 5. Отметим и повышение среднего объема клеток диатомей при N:P = 100. Цианобактерии имеют монотонную тенденцию к снижению среднего объема клеток по мере роста отношения азота к фосфору.
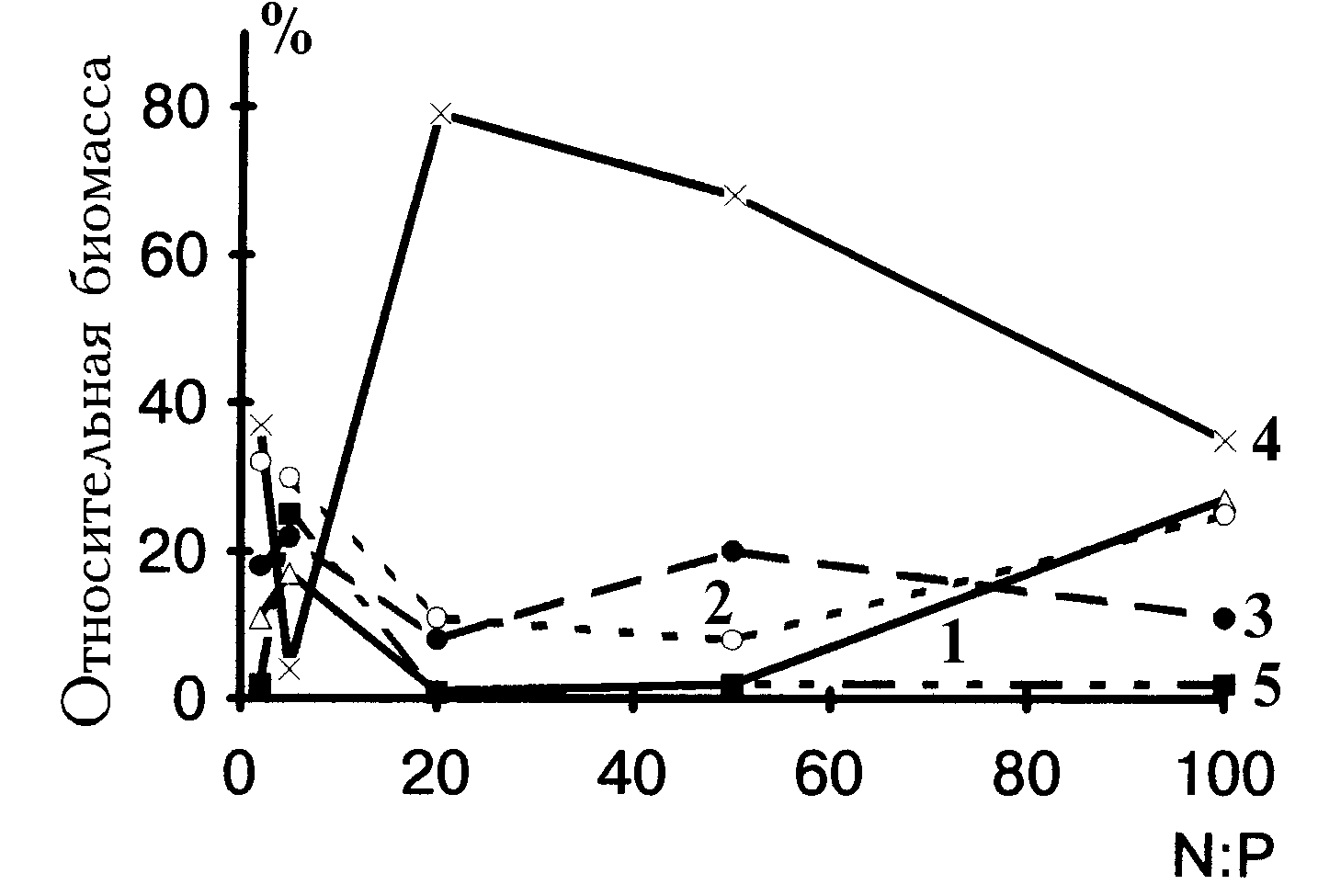
Относительные обилия размерных классов водорослей представлены на рис. 4.10. Поскольку клетки размером более 10 нг чрезвычайно редки в общей биомассе, к крупным клеткам в данном случае отнесены те, которые входили в диапазоны 1.1-3.2 и 3.3-10 нг. Так, особи из интервала 1.1-3.2 нг занимают доминирующее положение в сообществе при отношениях 20 и 50, при более низких и высоких отношениях их относительная биомасса понижается. Клетки массой 3.3-10 нг наиболее обильны при отношении, равном 5. Представительность трех самых мелких размерных классов падает при переходе от отношения 2 к отношению 50, однако при отношении, равном 100, они вновь занимают доминирующее положение.
Таким образом, отношение концентраций растворенных в воде азота и фосфора выступает в качестве одного из регулирующих факторов и для структуры прудового альгоценоза. Поместив фитопланктон в частично контролируемые условия, мы получили возможность следить непосредственно за потреблением и ростом клеток в той среде, которую сами создали. Результаты первого из описанных опытов свидетельствуют о том, что абсолютная концентрация фосфатов в исследованном диапазоне и на фоне заданных азотных добавок не влияет на микросукцессию в аквариумах. Изменение же содержания азотных солей действует примерно в том же направлении, что и изменение соотношения между этими элементами. Однако увеличение исходной концентрации азота в воде не всегда ведет к очевидным эффектам. Скажем, доминирующие виды протококковых при высоких концентрациях понижают свою конечную биомассу, т. е. при не слишком больших перепадах исходных концентраций азота отсутствует монотонность отклика. Кроме того, неочевидно также влияние азота на размерную структуру сообщества. Поэтому, с нашей точки зрения, распределение групп водорослей определяет все же отношение основных биогенных элементов. Но варьировать это соотношение, имея перед собой цель управления сезонной сукцессией, следует при помощи увеличения или уменьшения азотных добавок.
Что касается действия конкретных значений установленного фактора, то соотношения 15 и 20 наиболее благоприятно влияют на рост зеленых и, в частности, протококковых видов. Заметим, что и более высокие отношения (50 и 100) хотя и не приводят к абсолютным максимумам биомассы зеленых, но тем не менее никак не меняют их доминирующее положение в сообществе. Успешное развитие сине-зеленых определяется низкими отношениями азота к фосфору (2-5). Во всех остальных случаях рост отдела и составляющих доминантных видов заметно замедляется. Для диатомей, возможно, стимулирующими являются отношения, лежащие в пределах от 5 до 20.
Как было сказано выше, содержание биогенных веществ в питательной среде должно соответствовать потребностям в них клеток фитопланктона. Это касается как абсолютных концентраций азота и фосфора, так и их соотношения. Потребности фитопланктонных организмов в компонентах минерального питания (или клеточные квоты) сильно варьируют в зависимости от фазы роста (см. гл. 3). Согласно исходной гипотезе оптимальным для данного вида или группы видов водорослей является соотношение лимитирующих субстратов в воде, равное соотношению минимальных квот по ним для этих видов.
В гл. 3 содержится описание методов определения потребностей микроводорослей в азоте и фосфоре (Левич и др., 1986б; Левич, 1989). Там же приведены конкретные значения клеточных потребностей для ряда видов зеленых водорослей и цианобактерий, выращиваемых в лаборатории в условиях накопительного культивирования. Сопоставляя эти данные с конечными биомассами видов в прудовой поликультуре, приходим к выводу, что в среднем для зеленых водорослей соотношение потребностей близко к их оптимальному сочетанию в исходной среде, т. е. к 20. По-видимому, для некоторых видов протококковых стимулирующими могут выступить и более высокие отношения — порядка 30-40. Для
S. quadricauda соотношение биогенных элементов, приведшее к максимальному росту, оказалось в три раза меньшим, чем соотношение квот. В данном случае, скорее всего, в опыте по вычислению потребностей минимальная квота по азоту не была достигнута, а прекращение деления возникло в результате самозатенения клеток.Цианобактерии (
Anabaena, Anacystis) также могут обладать высоким отношением минимальных квот (около 20), тогда как рост представителей этого отдела в прудовом сообществе сдерживался именно этим отношением. Однако среди сине-зеленых доминантов пруда, эффективно развивавшихся при отношениях, равных 25, не было представителей указанных родов. По-видимому, преобладавший среди сине-зеленых Microcystis sp. (во втором опыте) имеет иной оптимум отношения N:P, что подтверждается данными Г.-Ю. Ри и Дж. Готама (Rhee, Gotham, 1980). Согласно этим данным, оптимальным для рода Microcystis является отношение азота к фосфору, равное 9. Эта величина выведена авторами из отношения потребностей водоросли в биогенных элементах и близка к оптимуму в наших опытах. Потребности водорослей в названной работе не совпадают с минимальными квотами, тем не менее соответствие между потребностями Microcystis и оптимальными для него дозами минеральных добавок говорит о неслучайности полученного результата. Данный пример характеризует цианобактерии как сильно эврибионтные организмы по отношению к сочетанию азота и фосфора: верхняя граница соотношения потребностей у них доходит до 100, как у Anabaena cylindrica (Dauta, 1982б).Изложенные результаты показывают, что путем варьирования количества и отношения вносимых азотных и фосфорных компонентов минерального питания возможно регулировать таксономический и размерный состав природного фитопланктона in vitro, и управление идет, по всей видимости, на уровне родов и даже видов водорослей. Для понимания механизмов, действующих при управлении на альгоценоз, и для проверки практической действенности метода необходимы эксперименты с лабораторными альгоценозами (см. 4.1) и испытания в природных водоемах (см. 4.3).
4.3. Натурные эксперименты
Еще в 30-е годы В.Пирсолл (Pearsall, 1930, 1932) одним из первых наблюдал, что различные отношения химических элементов, включая N:P, Ca
Mg : Na K, способствуют образованию различных сообществ фитопланктона в природе.В.Смит (Smith, 1983), проанализировав ситуацию по 12 озерам мира, обнаружил преобладание цианобактерий в период, когда величина отношения азота к фосфору имеет значе-ния < 25 (рис. 4.11). При атомном отношении N:P больше 25 преобладают зеленые и диатомовые микроводоросли. В некоторых из исследованных озер увеличение данного отношения произошло из-за отвода фосфорсодержащих сточных вод, при этом также возросла доля нефиксирующих азот зеленых и диатомовых водорослей. Д.Тилман (Tilman, 1982) называет закономерность, обнаруженную В.Смитом, “драматическим воздействием показателя отношения азота к фосфору на таксономический состав альгоценозов озер”.
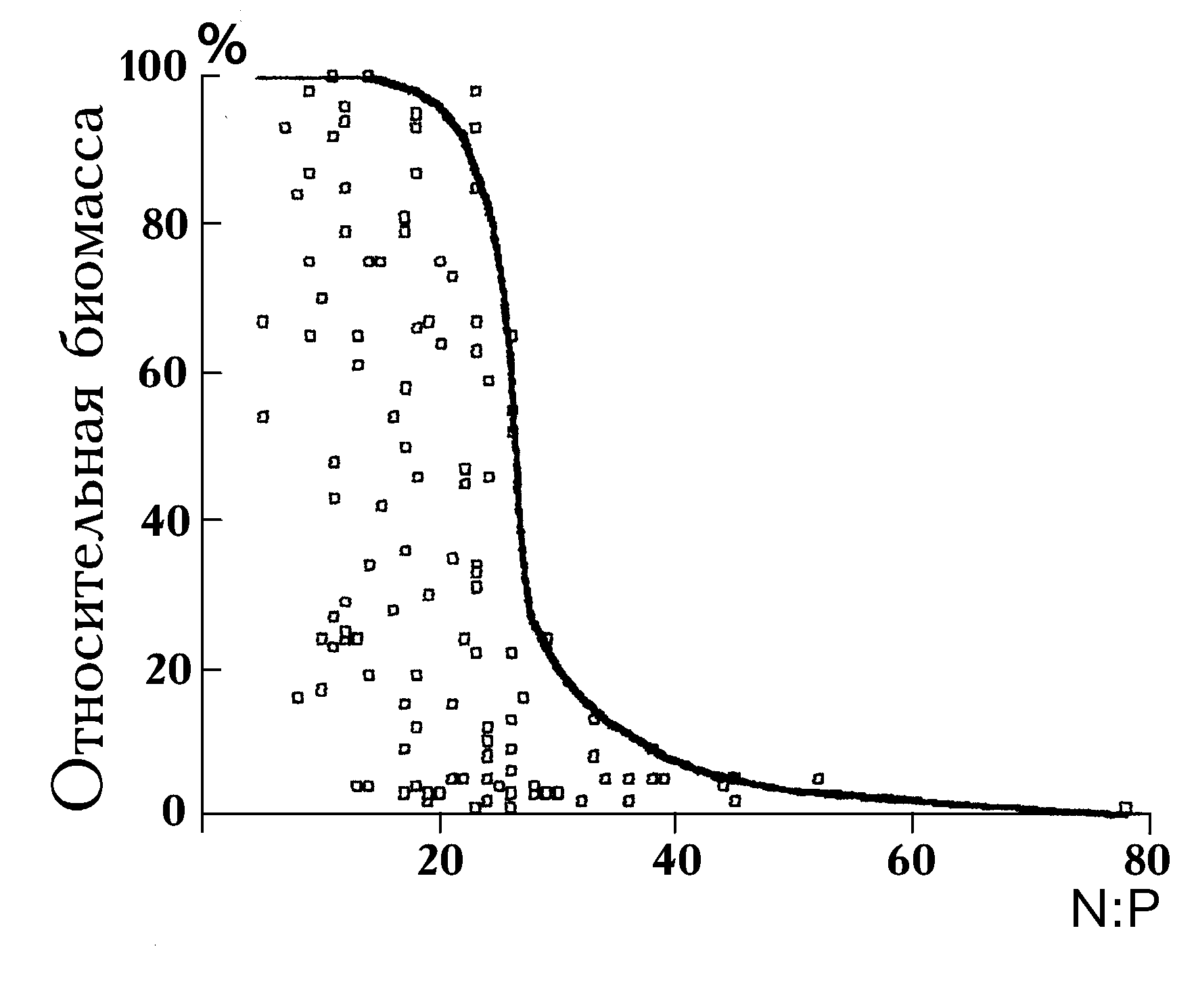
Позднее В.Смит (Smith, 1986) обобщил материалы уже по 22 озерам, включив в анализ условия освещения и морфометрию водоемов. Учет помимо отношения азота к фосфору дополнительных факторов позволил оставить в силе вывод о существенном влиянии отношения биогенных элементов на видовую структуру планктона.
Ф.Пик и Д.Лин (Pick, Lean, 1987), проанализировав ряд исследований по влиянию различных отношений питательных веществ на цианобактерии, пришли к выводу, что помимо высокой температуры, относительной стабильности водной толщи, концентраций углерода и железа, для этой группы водорослей также важны определенные отношения азота к фосфору и определенные световые условия.
Д.Шиндлер (Schindler, 1977) провел многолетнее экспериментальное изучение удобряемых малых озер. В одном из озер, куда в течение шести лет подряд добавляли удобрения с атомным отношением N:P, равным 30, на протяжении всего опыта в фитопланктоне доминировали водоросли рода
Scenedesmus. После этого пропорция азота и фосфора в удобрении в этом озере была снижена до 11 и в нем стали преобладать сине-зеленые азотфиксаторы рода Anabaena. В дальнейшем, когда и в первом озере отношение понизили до 5, в нём сменился тип “цветения”: доминирующее положение также заняли сине-зеленые (Aphanizomenon gracile). Д.Шиндлер подчеркивает, что сине-зеленые водоросли не только преобладают при низких отношениях азота к фосфору в неудобряемой воде исследованных озер, но их доминирование также может быть спровоцировано путем экспериментального внесения подкормки в указанных отношениях. Так, при снижении в удобрениях отношения азота к фосфору до 5 оказалось, что прирост биомассы фитопланктона в экспериментальном озере (в 48 раз по сравнению с неудобряемым) происходит почти полностью за счет сине-зеленых (Findley, Kasian, 1987). К сходному выводу привели исследования в озере Сойанг (Корея), где рост относительного обилия синезеленых водорослей произошел после постепенного увеличения с 1984 по 1989 гг. концентрации P в воде (на фоне большого количества N) и соответствующего уменьшения N:P от 100 до 50 (Cho et al., 1990). Обратный эффект наблюдался в южноафриканском водохранилище Хартбиспоорт (Haarhoff et al., 1992). Меры по очистке воды от хозяйственных стоков привели здесь к тому, что значение N:P в воде увеличилось от 4 до 25. Общая биомасса фитопланктона после этого снизилась, а ранее доминировавшие синезеленые водоросли (Microcystis aeruginosa) сменились зелеными. Обратная зависимость между численностью синезеленых водорослей и величиной N:P в озерных экосистемах доказана путем статистического корреляционного анализа (Varis, 1991).Существуют данные (Blomqvist
et al., 1989; Klapwijk, 1990), свидетельствующие о существенном изменении структуры альгоценоза при смене соотношения N:P в воде. Так, после внесения в шведское озеро Ньюпфатет (фоновое содержание общего N — 200 мкг/л, общего P — 4 мкг/л; N:P = 50) 100 мкг N и 10 мкг P на 1 л (N:P = 10) значительно увеличилась биомасса Merismopedia tenuissima, Peridinium inconspicuum, Dyctiosphaerium botritella, хотя до внесения добавок доминировал только первый из этих видов (Blomqvist et al., 1989). Соотношение неорганических форм N и P в бассейне р.Рейн за последние 45 лет сильно уменьшилось (Klapwijk, 1990). Это привело к тому, что за эти годы видовой состав фитопланктона стал беднее, так как несколько таксонов исчезли, а численность других многократно уменьшилась.Зависимость обилия цианобактерий от низких отношений азота к фосфору и от достаточного снабжения фосфором была получена также при изучении “цветения” фитопланктона в озере Кеннеди (Stockner, Shortreed, 1988).
Если до сих пор речь шла о влиянии на структуру фитопланктона отношения минеральных форм азота и фосфора, то, согласно Д.Маккуину и Д.Лину (McQueen, Lean, 1987), рост усредненного годового процента цианобактерий стимулировался уменьшением отношения нитратного азота к общему фосфору. И, наоборот, когда это отношение было больше 5, никогда не наблюдалось массового развития сине-зеленых.
На состав природного альгоценоза также влияет взаимодействие N, P и Si. Долговременные изменения гидрохимических показателей дельты р.Рейн, выразившиеся в снижении величин Si:N и Si:P, способствовали массовому развитию динофлагеллят и синезеленых водорослей (Admiraal, Vlugt, 1990). Избавиться от этого явления можно за счет снижения азотной и фосфорной нагрузки.
Дж.Шеймесс с соавторами (Shamess
et al., 1990) пришли к выводу, что регуляторами цветения фитопланктона могут быть соотношения не только N, P и Si, но и некоторых других веществ и даже микроэлементов. Так, в их опытах соотношение концентраций солей серной кислоты и Mo5+ определяло доминирующую роль в сообществе азотфиксирующих синезеленых водорослей. Авторы подчеркивают, что абсолютные концентрации этих ионов не влияли на процесс фиксации N.Таким образом, отношение концентраций в воде незаменимых химических элементов (азота и фосфора) признается многими исследователями фактором, прямо влияющим на соотношение биомасс различных водорослей в естественном сообществе.
По мнению ряда авторов (Rhee, Gotham, 1980; Tilman, 1981; Terry, 1985), оптимальным считается такое начальное отношение компонентов среды, при котором прирост биомассы после достижения стационарной фазы роста культурой возможен только при одновременном добавлении всех элементов и не происходит при добавлении лишь части из них. Иначе говоря, оптимальность понимается в смысле сбалансированного (полного) потребления начальных количеств всех субстратов. Если в такой ситуации вещества потребляются полностью, то оптимальное отношение их начальных концентраций в среде должно оказаться равным отношению соответствующих минимальных квот для данного вида.
Имевшиеся в распоряжении авторов литературные и оригинальные данные позволили определить тенденции в оптимальности отношения азота к фосфору для крупных таксонов микроводорослей. Высокие отношения N:P (более 20) в основном благоприятны для зеленых водорослей, в частности для представителей массового рода
Scenedesmus. В то же время для многих видов диатомей характерны оптимальные отношения в диапазоне 7-12, а для синезеленых — в пределах от 4 до 10, за исключением ряда таксонов, например родов Anacystis (Сиренко, 1972) и Synechococcus (Suttle, Harrison, 1988), обладающих высокими стимулирующими отношениями N:P.