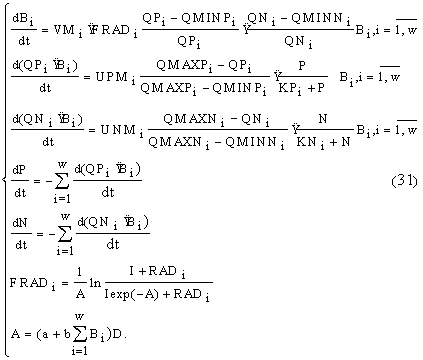
Глава 5
. Имитационное моделированиеКультивирование водорослей в лабораторном альгоценозе возможно при постоянных температуре, освещенности, практическом отсутствии пространственной неоднородности, отсутствии консументов, редуцентов, а также при строгом контроле поступающих форм биогенных элементов (2.1). Благодаря этому обеспечивается адекватность эмпирического материала и
модели, учитывающей только немногие из факторов, влияющих на рост фитопланктона. Такая модель может быть тщательно откалибрована и верифицирована и, кроме того, позволяет достаточно детально изучить влияние этих немногих факторов, в частности, исследовать закономерности смены доминирующих видов в зависимости от различных соотношений биогенных элементов. Возможность охвата широкого диапазона соотношений помогает выявить наиболее перспективные с точки зрения управления соотношения без проведения многочисленных натурных экспериментов. Возможность проведения необходимых экспериментов с лабораторным альгоценозом облегчает проверку предсказательной силы модели.Разработанная модель была использована для исследования влияния различных концентраций азота и фосфора в среде на структуру лабораторного водорослевого сообщества (Левич, Личман, 1992). Остановимся подробнее на рассмотрении имитационной модели.
Формулировка модели. Модель построена для лабораторного альгоценоза, функционирующего в условиях накопительного культивирования.
Конкретный вид уравнений определяется возможностью содержательной интерпретации, степенью адекватности описания реальных данных, относительной простотой.
Процессы роста и поглощения описываются раздельно. Скорость роста биомассы изменяется гиперболически и, согласно современным представлениям (Йоргенсен, 1985), зависит от внутренних концентраций биогенных элементов (модель Друпа). Скорость поглощения каждого биогенного элемента из среды в рассматриваемых условиях предполагается равной скорости изменения общего внутриклеточного запаса данного элемента и определяется как его внутриклеточным, так и внешним уровнем.
Помимо динамики биомассы фитопланктона и внутриклеточного содержания биогенных элементов (азота и фосфора), моделируется динамика концентрации азота и фосфора в среде. В соответствии с законом сохранения вещества, применительно к конкретным условиям, скорость изменения концентрации биогенного элемента в среде по абсолютной величине равна скорости поглощения этого элемента фитопланктоном.
Таким образом, для каждого вида в модель включено три уравнения: динамики биомассы и динамики внутриклеточного содержания азота и фосфора, а для всего сообщества: уравнения динамики концентраций азота и фосфора в среде:
Переменные состояния модели: B
i — концентрация биомассы i-го вида; QPi — клеточная квота в единице биомассы по фосфору; QNi — клеточная квота в единице биомассы по азоту; N и P — концентрации азота и фосфора в среде.Параметры модели: VM
i — максимальная удельная скорость роста i-го вида; UPMi и UNMi — максимальные удельные скорости поглощения фосфора и азота i-м видом; QMINPi, QMAXPi и QMINNi, QMAXNi — минимальная и максимальная клеточные квоты по фосфору и азоту соответственно; KPi, KNi — константы полунасыщения по фосфору и азоту для i-го вида; RADi — константа радиации для i-го вида; I — уровень освещенности; a — коэффициент поглощения света водой; b — коэффициент поглощения света фитопланктоном; D — глубина, w — количество видов.Помимо прямого учета моделью конкуренции за биогенные элементы и конкурентного взаимодействия путем затенения, можно предложить косвенный учет в модели других взаимодействий видов, например, посредством метаболитов, играющих, по-видимому, важную роль в сообществах фитопланктона (Алексеев, 1976; Федоров, Кафар-Заде, 1976; Страшкраба, Гнаук, 1989). Возможный путь такого учета несубстратных взаимодействий видов в данной модели — зависимость физиологических параметров видов от состава сообщества, на возможность существования которой указывается в ряде публикаций (Страшкраба, Гнаук, 1989; Knoechel, Kalff, 1978; J
Ш rgensen, 1988).При плохой изученности несубстратных взаимодействий видов, при отсутствии приемлемых способов их математического описания нахождение конкретных значений параметров для альгоценозов определенного состава путем калибровки может рассматриваться как способ учета в модели этих взаимодействий. В дальнейшем, по мере накопления более полной информации, безусловно необходимо будет разрабатывать прямые способы описания несубстратных взаимодействий видов конкретными функциональными зависимостями.
Версия модели, при которой параметры вида могут меняться в зависимости от присутствия в сообществе тех или иных видов, с одной стороны, вероятно, увеличивает реалистичность модели, а с другой — затрудняет ее применение, уменьшая универсальность. Поэтому гипотеза о том, что значения параметров видов практически не зависят от состава сообщества, т.е. являются константами, представляется более удобной, но
также нуждается в проверке.Анализ чувствительности модели. Был проведен анализ чувствительности моделей четырех видов монокультур из опыта №7 (Левич, Личман, 1992):
Scenedesmus quadricauda (Turp.) Breb., S. obliquus (Turp.) Kutz., Ankistrodesmus falcatus (Corda) Ralfs. и Chlorella vulgaris Beijer. и модели их четырехвидовой поликультуры. Конкретные значения параметров (табл. 5.1), в области которых исследовалась чувствительность, были получены калибровкой по данным монокультур.Производилось варьирование значений каждого из параметров в отдельности на 5, 10, 20 % при неизменных остальных параметрах. Кроме того, варьировались по отдельности начальные значения переменных состояния модели. Для оценки чувствительности модели к изменению параметров и начальных значений переменных было выбрано несколько показателей,
исчерпывающе характеризующих поведение модели. Для моделируемых динамик биомасс в моделях монокультур это: биомасса вида после выхода в стационарное состояние; время выхода биомассы в стационарное состояние; тангенс угла наклона кривой динамики биомассы, определявшийся как разность между стационарной и начальной биомассой, отнесенная ко времени выхода биомассы в стационарное состояние. В модели поликультур: величина суммарной биомассы сообщества после достижения стационарной фазы роста всеми видами; время достижения суммарной биомассой сообщества стационарной фазы; биомасса каждого вида в момент остановки роста. Для динамик концентраций азота и фосфора в среде: время исчерпания биогенного элемента в среде; начальная концентрация биогенного элемента в среде, отнесенная ко времени его исчерпания. Анализировалась величина отклонения (в %) значения показателя по сравнению с таковым при неизменных параметрах и фиксированных начальных значениях переменных состояния.
Из проведенного анализа чувствительности модели монокультур следует, что параметрами, изменение которых оказывает наиболее сильное влияние на характеристики модели, являются удельная максимальная скорость роста, минимальные квоты по лимитирующему биогенному элементу и видоспецифическая константа радиации. Поэтому, как правило, имеет смысл затрачивать усилия в процессе разработки модели на уточнение именно этих параметров (например, методом автоматизированной калибровки, а, возможно, и в ходе лабораторных экспериментов).
Анализ чувствительности модели к изменению начальных значений переменных состояния показывает, что начальные концентрации азота и фосфора в среде должны быть определены наиболее точно, так как уже двадцатипроцентное изменение сильно влияет на выходные характеристики модели, тогда как изменение начальных значений биомасс и освещенности на 20% (а биомасс даже в два раза) и начальных квот в пять раз существенно не отражается на поведении модели монокультур.
Анализ чувствительности модели поликультуры выявля- ет, что наибольшее влияние на выбранные показатели оказывает изменение параметров вида с наименьшей минимальной квотой (сравнительно с другими видами) по лимитирующему биогенному элементу, высокой удельной максимальной скоростью роста и не самой маленькой начальной биомассой (в нашем случае таким видом является
S. quadricauda ).В отличие от модели монокультур, модель четырехвидовой поликультуры оказалась чувствительной к изменению начальных биомасс видов. Изменение начальных квот видов на полпорядка и на порядок также повлекло изменение значений выходных характеристик модели. На выбранные показатели модели поликультур заметно влияли изменения начальных концентраций азота и фосфора, а также уровня освещенности.
Проведенный анализ чувствительности модели поликультуры показывает, что наряду с калибровкой удельной максимальной скорости роста, минимальных квот по фосфору и азоту и видоспецифической константы освещенности желательна калибровка значения скорости поглощения биогенного элемента из среды. Кроме того, модель поликультуры нуждается в большей по сравнению с моделью монокультуры точности определения значений начальных биомасс и начальных квот.
Необходимо отметить, что возможная неодинаковая чувствительность модели в разных областях пространства параметров затрудняет прямое использование результатов анализа чувствительности для отбора параметров, нуждающихся в уточнении.
Калибровка и верификация модели. С целью получения необходимых для калибровки и верификации данных был поставлен эксперимент с лабораторными альгоценозами из одного и нескольких видов пресноводных микроводорослей (см. описание опыта №10 в разделе 2.3).
Так как наблюдался разброс значений численности, а следовательно и биомасс, обусловленный ошибкой измерений, для более эффективного использования экспериментальных данных при разработке модели была проведена рекомендуемая при моделировании первичная статистическая обработка данных — сглаживание кривых динамик биомасс. Сглаживание проводилось с помощью линейных интерполяционных полиномов.
В данной работе критерием качества калибровки и верификации служил коэффициент несовпадения Тейла (Тейл, 1971) в следующем виде:
 , (32)
, (32)
где P
i — прогнозируемая величина; Ai — соответствующий действительный результат; N — число проверяемых значений.Пороговое значение выбранного критерия, т.е. максимальное значение, при котором предсказания модели считались приемлемыми, определялось с учетом ошибок измерения биомасс. В каждом конкретном случае калибровки (калибровка монокультур, двухвидовых поликультур, трехвидовых поликультур) критериальные ограничения несколько варьировали, но не превышали 0.45 для критерия Е (что соответствует ошибке измерения, не превышающей 50%). Пробные точки из пространства параметров, удовлетворяющие имеющемуся критериальному ограничению, оставлялись. Соответствующие этим точкам сочетания параметров затем верифицировались на других данных.
Модель монокультур. Для каждого вида одновременно калибровались следующие параметры: максимальная удельная скорость роста, минимальные квоты по фосфору и азоту, видоспецифическая константа радиации, максимальные квоты по фосфору и азоту, максимальные скорости поглощения фосфора и азота, а также начальные квоты по этим биогенным элементам. Способ включения этих параметров в модель дан в системе уравнений (31). Значения для констант (а
= 0.27, b = 0.18, KN = 4.0, KP = = 0.1) были взяты из литературы (Йоргенсен, 1985). Константы полунасыщения по азоту и фосфору не калибровались, так как к их изменению модель была нечувствительной.Для всех трех видов калибровка проводилась на данных динамик биомасс на среде 1 (табл. 5.2). Приблизительный диапазон возможных изменений параметров был взят из литературы (Banse, 1982; Йоргенсен, 1985; J
Ж rgensen, 1988).Было просмотрено 600 пробных точек в пространстве параметров. Удачные наборы параметров, найденные калибровкой, верифицировались на данных других сред. Были оставлены только те наборы параметров, для которых коэффициент Е (см. формулу 32) не превышал 0.3 (табл. 5.3). Затем эти наборы параметров проверялись на данных, полученных после пересева монокультуры с каждой среды на две. Наблюдалось хорошее согласование модельных и экспериментальных динамик. Найденные значения для максимальных скоростей роста и поглощения азота и фосфора очень близки к значениям, найденным экспериментально (Левич, Артюхова, 1991).
Модель двухвидовых поликультур. Для модели двухвидовой поликультуры калибровались те же параметры, что и для монокультур, но уже одновременно для двух видов. Калибровка проводилась по данным динамик биомасс на среде 2 (табл. 5.2). Найденные наборы параметров верифицировались по данным динамик биомасс на среде 4 (табл. 5.2). Оставлялись наборы параметров, с которыми коэффициент E не превышал 0.35 (табл. 5.3).
Графически также наблюдалось хорошее согласование модельных и экспериментальных динамик био-масс (см. на рис. 5.1 динамики биомасс, предсказанные моделью с одним из найденных наборов, и экспериментальные динамики).
Для S. quadricau- da калибровка про-водилась по данным поликультуры S. quad-ricauda и A. variabi- lis. Найденные параметры верифицировались по данным поликультуры этого вида с A. falcatus, для A. falcatus калибровка проводилась по данным поликультуры этого вида с A. variabilis. Полученные параметры проверялись на данных поликультуры A. falcatus и S. quadricauda. Для вида A. variabilis в табл. 5.3 объединены параметры, полученные в результате этих двух калибровок.
Предсказания модели двухвидовых поликультур с откалиброванными и верифицированными наборами параметров хорошо согласовывались с экспериментальными данными, полученными пересевом каждой поликультуры со всех сред на две среды (аналогично монокультурам).
Модель трехвидовой поликультуры. Калибровались те же параметры, что и для монокультур и двухвидовых поликультур, для трех видов одновременно. Для калибровки использовались данные динамик биомасс на среде 2 (см. табл. 5.2), для верификации — данные со сред 2, 3 и 4 (см. табл. 5.2). Наборы параметров считались приемлемыми, если при верификации критерий Е также не превышал 0.45 для A. variabilis, и 0.35 для S. quadricauda, 0.30 — для A. falcatus. Значения параметров и критерия Е см. в табл.5.3.
Сопоставление предсказаний откалиброванной модели и данных динамик биомасс после пересева трехвидовой поликультуры со всех сред на две не выявило противоречий в семи из восьми случаев.
С целью проверки гипотезы о том, что значения параметров каждого вида не зависят от состава сообщества, т.е. являются константами в рассматриваемой ситуации, наборы параметров, откалиброванные и верифицированные на монокультурах, были подставлены в модели для двух- и трехвидовой поликультур. Кроме того, в модель трехвидовой поликультуры были подставлены еще и параметры, полученные в результате калибровки и верификации по данным двухвидовых поликультур, а параметры, откалиброванные по трехвидовой поликультуре, были подставлены в модель монокультур и двухвидо- вых поликультур. Наблюдались значительные расхождения между моделируемыми и экспериментальными значениями биомасс (значения критерия Е (формула (32)) превышали порого- вое значение 0.45, вычисленное с учетом 50%-й ошибки).
Таким образом, оказалось, что модель с параметрами, откалиброванными и верифицированными по данным монокультур, не может удовлетворительно описать динамики биомасс в поликультурах, и модель с параметрами, полученными аналогично для двухвидовых поликультур, не описывает трехвидовую поликультуру. Для видов S. quadricauda и A. variabilis в поликультурах наблюдался меньший рост, чем предсказывался моделью с параметрами монокультур, а для трехвидовой поликультуры и с параметрами, полученными для двухвидовых культур. Для вида A. falcatus обнаружилось, что некоторые параметры, полученные для него по данным монокультуры, при подстановке в модель поликультур приемлемо описывали динамику биомассы в поликультурах. Параметры, найденные калибровкой и верифицированные по данным трехвидовой поликультуры, неудовлетворительно описывали динамики видов в моно- и двухвидовых поликультурах.
В результате проделанного анализа, включавшего также сопоставление значений параметров, полученных в ходе различных калибровок, был сделан вывод о зависимости значений параметров видов от состава сообщества. Наблюдалось, в частности, увеличение значений минимальных квот видов S. quadricauda и A. variabilis при переходе от монокультур к двухвидовым и трехвидовым поликультурам. Данная зависимость значений параметров от состава сообщества указывает, по-видимому, на существование каких-то, возможно, метаболитных, взаимодействий видов, не включенных в модель непосредственно.
Таким образом, для каждого вида имеется три совокупности наборов параметров: калибровавшиеся и верифицировавшиеся по данным монокультур, двухвидовых поликультур и трехвидовых поликультур (см. табл. 5.3).
Что касается приемлемости модели, возможности ее использования, то на основе результатов, полученных из имеющихся в нашем распоряжении данных, можно сделать вывод, что модель является приемлемой при условии существования зависимости значений параметров от состава сообщества и может быть использована с наборами параметров, найденными для конкретного состава сообщества.
Модельный анализ влияния различных концентраций биогенных элементов на структуру альгоценоза. С разработанной имитационной моделью были проведены машинные эксперименты по выявлению влияния различных комбинаций начальных концентраций азота и фосфора в среде на обилие видов в сообществе.
В опыте №7 проводился анализ структуры четырехвидового сообщества микроводорослей в зависимости от начальных концентраций азота и фосфора в среде. Определение биомасс проводилось на момент достижения всеми видами стационарной фазы роста. Конкретные значения параметров были получены калибровкой по монокультурам видов S. quadricauda, S. obliquus, A. falcatus и Ch. vulgaris (см. табл. 5.1).
Нижняя граница рассматриваемого диапазона начальных концентраций в среде определялась совокупным для всех видов внутриклеточным содержанием азота и фосфора. Концентрации ниже этого значения не рассматривались, так как они не оказывали бы существенного влияния на структуру сообщества. Верхний предел концентраций был задан, исходя из информации об отсутствии токсического влияния данных концентраций на большинство видов.
В первом машинном эксперименте было испытано 45 различных комбинаций начальных концентраций азота и фосфора в среде. Концентрация азота варьировала от 0.5 до 30 мг/л, концентрация фосфора — от 0.2 до 100 мг/л. Наблюдалась закономерная смена доминирующих видов при изменении соотношенийа азота к фосфору, объясняющаяся видовыми различиями численных значений таких параметров, как максимальная скорость роста биомассы, максимальные скорости поглощения азота и фосфора, минимальные квоты по этим биогенным элементам.
Результаты первого машинного эксперимента были сгруппированы по характеру распределения относительных биомасс видов в зависимости от соотношения суммарного (клеточные запасы плюс содержание в среде) азота к суммарному фосфору (табл.5.4). Кроме того, для каждого вида был определен лимитирующий рост биогенный элемент путем сравнения конечной клеточной квоты с минимальной клеточной квотой по данному элементу. Для элемента, обусловившего остановку роста биомассы, значения этих величин очень близки.
Благодаря наименьшей из всех видов минимальной клеточной квоте по азоту и максимальной скорости поглощения азота S. quadricauda получает конкурентное преимущество в утилизации данного ресурса в областях низких и средних концентраций азота (малые соотношения азота к фосфору), где происходит лимитирование этим биогенным элементом, приводящее к наибольшей доле S. quadricauda в сообществе.
С увеличением отношения N:P снижается доля S. quadricauda за счет вытеснения его A. falcatus. A. falcatus — единственный вид, чей рост во всем диапазоне начальных концентраций азота и фосфора лимитируется только азотом за счет самой маленькой по сравнению с остальными видами минимальной квоты по фосфору.
Интересно, что максимальных относительных биомасс эти виды достигают при значениях N:P, близких к отношению минимальных квот по азоту и фосфору (QMINN:QMINP) для данного вида. По мере удаления величин N:P от этого значения происходит уменьшение доли вида в сообществе.
S. obliquus
имеет соотношение минимальных квот, близкое к таковому S. quadricauda, но не достигает высоких значений биомассы при благоприятных значениях отношения N:P (близких к соотношению минимальных квот). Это объ-ясняется тем, что такие его параметры, как максимальная скорость роста, поглощение азота, а также начальная биомасса, имеют наименьшие значения.Ch. vulgaris
занимает промежуточное положение по ряду параметров по сравнению с другими видами. За счет этого она не становится доминирующим видом, однако при отношени- ях азота к фосфору, близких к отношению ее минимальных квот, достигает своей максимальной, наблюдаемой в машинном эксперименте, биомассы.С целью более подробного анализа влияния различных соотношений концентраций азота и фосфора был проведен второй машинный эксперимент. Относительные биомассы ви- дов определялись для 240 комбинаций начальных концен-траций азота и фосфора в среде.
Рассмотрен более широкий диапазон изменений отношения N:P: от 0.07 до 220. Результаты этого эксперимента подтверждают закономерности смены доминирующих видов в зависимости от N:P, обнаруженные в предыдущем эксперименте (табл. 5.5).
Закономерное изменение доли видов в зависимости от отношения начальных концентраций суммарного азота и фосфора происходит как при изменении концентраций азота и фиксированной концентрации фосфора, так и наоборот, при неизменной концентрации азота и изменении концентраций фосфора (рис. 5.2).
Наблюдаемая в машинном эксперименте смена доминирующих видов в зависимости от соотношения азота и фосфора, обусловленная видовыми различиями в способности утилизировать ресурс, свидетельствует о принципиальной возможности управления структурой сообщества путем изменения отношений азота к фосфору.
Данные выводы были проверены на модели другой поликультуры, состоящей из трех видов (опыт №10):
S. quadricauda, A. falcatus и A. variabilis. С этой целью был проведен третий машинный эксперимент. Конкретные значения параметров (табл. 5.3) были получены калибровкой и верифицированы на экспериментальных данных по трехвидовой поликультуре. В этом эксперименте анализировали диапазон изменений отношений азота кфосфору от 0.1 до 100 (всего 17 отношений). Причем в одном варианте машинного эксперимента изменяли отношение азота к фосфору за счет изменения концентрации фосфора при фиксированной концентрации азота в среде (таких концентраций азота было восемь: от 2 мг/л до 40 мг/л). Во втором варианте при каждой из восьми различных концентраций фосфора (от 0.1 до 15 мг/л) соотношение N:P изменяли варьированием концентрации азота.Относительные обилия видов анализировали на сороковые сутки моделируемого эксперимента. Обнаруженная для четырехвидовой поликультуры закономерность — увеличение относительного обилия вида по мере приближения соотношения начальных концентраций азота и фосфора к значениям, близким к соотношению минимальных квот данного вида, — была выявлена и в этом машинном эксперименте с трехвидовой поликультурой (рис. 5.3 для видов S. quadricauda и A. variabilis
).Однако для вида
A. falcatus при фиксированных концентрациях азота и при уменьшении концентрации фосфора эта зависимость не выражена: относительное обилие вида возрастает даже при удалении соотношения N:P от значений, близких к соотношению минимальных квот. Это, по-видимому, объясняется влиянием абсолютных значений минимальных квот. У указанного вида самая маленькая минимальная квота по фосфору и в условиях возрастания степени фосфорного лимитирования способность данного вида лучше утилизировать этот ресурс позволяет ему наращивать биомассу, тогда как другие виды делать это не в состоянии.Обнаружилось также, что при одних и тех же отношениях азота к фосфору, близких к отношению минимальных квот видов, но при увеличении абсолютных количеств биогенных элементов, возрастает относительное обилие видов
S. quadricauda и A. variabi-lis. Это происходит, по-видимому, в результате того, что при низких концентрациях фосфора вид A. falcatus имеет преимущество за счет самой маленькой минимальной квоты по фосфору, а при возрастании концентрации фосфора это преимущество оказывает меньшее влияние на исход конкуренции.На основании полученных результатов можно подтвердить эмпирический вывод о том, что существует реальная возможность управления структурой альгоценозов с помощью варьирования соотношений концентраций биогенных элементов, в частности азота к фосфору. Однако обнаруженная зависимость не абсолютна (она может проявляться в виде более или менее отчетливых тенденций, а может нивелироваться влиянием других факторов). Поэтому, несомненно, при попытке управления структурой альгоценозов необходимо учитывать остальные характеристики видов (помимо соотношения минимальных квот), так как они могут существенно влиять на конечное распределение относительных обилий видов в сообществе. И здесь модельный анализ может способствовать повышению эффективности управления структурой альгоценозов.