Часть II. Модели многовидовых сообществ фитопланктона
Во второй части пособия рассматриваются математические модели, которые позволяют выявить предполагаемые экологические механизмы, ответственные за рост микроводорослей и регулирование состава сообщества. Сравнение полученных модельных данных с результатами лабораторного эксперимента дает возможность оценить адекватность математической модели, ее числовые параметры, справедливость предсказываемых эффектов управления.
По отношению к вопросам управления экологическими сообществами выделены и обсуждаются два аспекта теоретического исследования:
4.1. Формула видовой структуры
Можно указать по крайней мере два пути получения уравнений динамики в теоретическом естествознании. Первый путь — постулирование уравнений. Например, уравнения Ньютона в механике или уравнение Вольтерра в динамике популяций. Второй путь — постулирование или вывод функционалов, для которых искомые уравнения получаются с помощью вариационных процедур в качестве уравнений Эйлера-Лагранжа. В настоящей главе описывается применение вариационного подхода к моделированию в экологии сообществ.
Экологическое сообщество моделируется с помощью структуры множеств с разбиениями (Левич, 1978; Левич, 1980). Элементы множеств — это особи, классы разбиения — популяции биологических видов. Переходы сообщества из одних состояний в другие описываются соответствиями между множествами с разбиениями. Обычно из постановки задачи ясно, какие допускаются соответствия. Например, если мы хотим описать смертность особей, то допускаются не всюду определенные соответствия из одного множества в другое; если мы описываем размножение особей, то рассматриваются многозначные (не функциональные) соответствия и т. д. С каждым состоянием сообщества, т.е. с соответствующим множеством с разбиением А, мы свяжем следующую величину, называемую энтропией состояния А:
![]() ,
,
где
Рассмотрим сообщество, состоящее из организмов одного трофического уровня без возрастной структуры (например, из клеток фитопланктона), принадлежащих w видам и потребляющих m взаимонезаменимых ресурсов. Пусть в рассматриваемом сообществе допустимы деление клеток и их гибель, но недопустимы поглощение одних организмов другими и интродукция особей извне. Таким требованиям удовлетворяют инъективные, сюръективные, не всюду определенные и не функциональные соответствия. Энтропийный функционал для такого сообщества приобретает вид (Левич, 1982; Левич, 1988):
 ,
,
где n
i — численность в сообществе организмов вида i; n обозначает совокупность (n1, n2, ..., nw) и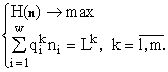
Здесь L
k — наличное количество k-го ресурса в среде;![]() ,
,
называемую формулой видовой структуры (Левич, 1980). Численности n
i и n оказываются функциями ресурсных потоков Lk. Эти функции отыскиваются как решения m алгебраических уравнений для множителей Лагранжа dk, зависящих от mw пaраметров — клеточных квотВ дальнейшем изложении изучается вариационная модель с ограничениями в виде неравенств, а не равенств, поскольку обычно известно, какой набор ресурсов необходим сообществу, но неизвестно, какие из этих ресурсов потребляются полностью.
Рассматривается задача на условный экстремум с ограничениями в виде неравенств (Левич и др., 1994):
 (4)
(4)
Здесь численности видов n
i переобозначены как xi, чтобы подчеркнуть приближение дискретных переменных ni действительными числами. Кроме того, всюду в дальнейшем, не оговаривая каждый раз особо, мы будем считать, что выполнены следующие естественные с точки зрения экологии требования:![]() .
.
От задачи (4) можно перейти к задаче
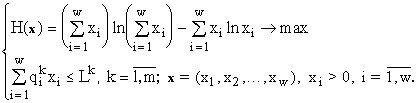 (5)
(5)
Гарантировано, что максимум энтропии Н в задаче (5) достигается.
Заметим, что (5) проще (4), так как на множестве x
i > 0,Необходимые условия экстремума. В соответствии с общим рецептом решения экстремальных задач с ограничениями в виде неравенств (Галеев, Тихомиров, 1980) методом Лагранжа выпишем необходимые условия экстремума в задаче (5):
 ;
;
![]() ; (6)
; (6)
![]() .
.
Из (А) мы получаем формулу видовой структуры (Левич, 1980):
![]() , (7)
, (7)
где
qi  (8)
(8)
Вычисляя из (8) n и
d, мы находим xi. Из первого уравнения (8) следует, что существует dj > 0,Из теоремы стратификации (см. 4.3) получаем, что для любого фиксированного вектора
L = (L1,L2,...,Lm) формула видовой структуры (7) получается из решения системы 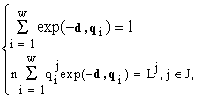 (9)
(9)
где J — некоторое подмножество индексов, JÌ
{1; 2; ...; m}.Инвариантность решений к преобразованию факторов среды.
Исключив n из (9), можно прийти к системе только для d  (10)
(10)
Из (10) мы видим, что компоненты
d зависят лишь от отношений ресурсных факторов, а так как относительные численностиИз системы (9) видно, что при таких преобразованиях
n® an, и, следовательно, xi® axi,Инвариантность решений к преобразованию потребностей видов.
Если мы теперь подвергнем аналогичным преобразованиям вектора квот qi® aqi,Зависимость полной численности и энтропии от факторов среды.
Из (10) мы видим, что = Lkf(v), где v имеет компонентами отношения ресурсных факторов, а f — некоторая функция. Из (10) мы видим, что v имеет компонентами
= Lkf(v), где v имеет компонентами отношения ресурсных факторов, а f — некоторая функция. Из (10) мы видим, что v имеет компонентами  , (11)
, (11)
и так как x
i = npi, то для H(x) =4.1.3. Примеры функции видовой структуры
Однофакторное лимитирование.
Обозначим символом d единственный множитель Лагранжа. Обозначим потребность i-го вида в единственном факторе через qi. Тогда система (8) приобретает вид:  (12)
(12)
В частности, мы еще раз видим. что при m
= 1 задачи с равенствами и неравенствами эквивалентны.Рассмотрим экспоненциальную модель распределения потребностей видов q
i=li. Пусть w >> 1. Тогда, заменяяТаким образом, при обоих приближениях качественное поведение функций оказывается одинаковым: Н и n пропорциональны L/
l, а ni — экспоненциально убывает.Рассмотрим теперь гиперболическую модель распределения потребностей видов: q
i=l(1+ln i). Снова, считая w >> 1 и переходя от суммирования к интегрированию, получаем n = (L/l) ((b-1)/b), H = b (L/l), ni= (L/l) ((b-1)/b) exp {-b(1 + ln i)}, где b — такое число, что (b-1)expb = 1 (b » 1.28; (b-1)/b » 0.23).Таким образом, снова видим, что H и n пропорциональны L/
l, а ni~i-b.Кроме того, при m
= 1 оказывается, что d = const и поэтому H = dL ~ L и n = ~ L. Тогда при m = 1 H~n. Однако уже при m = 2 существуют примеры, когда H и n не пропорциональны.
~ L. Тогда при m = 1 H~n. Однако уже при m = 2 существуют примеры, когда H и n не пропорциональны.
Двухфакторное лимитирование. Случай m
=2 для задачи с равенствами (9) был подробно изучен в одной из работ (Левич и др., 1993а), из результатов которой следует способ расчета конуса 
 . (13)
. (13)
В таком случае вектор
L = (L1, L2) лежит в конусеМожно также показать, что H(
L), xi(L), n(L), полученные при решении (4) при m = 2, зависят от L непрерывно,Рассмотрим некоторые конкретные примеры при
m=2. Используем собственные экспериментальные данные o потребностях в азоте (N) и фосфоре (P) четырех видов протококковых микроводорослей (размерность потребностей 10-9 мг/кл):![]()
Коэффициенты v
0 и v1, определяющие границы областей на рис. 4.1, равны v0 = 3.06 и v1 = 4.36. Результаты, полученные при численном исследовании задачи, представлены на рис. 4.2, 4.3, 4.4.Пусть теперь
 (14)
(14)
Cлучаи, когда d
1, d2 равны 0, оказываются невозможными, так как интегралы расходятся, и поэтому d1 > 0, d2 > 0. В таком случае наша задача сводится к задаче с равенствами. Некоторые результаты численного исследования (14) приведены на рис. 4.5, 4.6, 4.7.Трехфакторное лимитирование. Пусть w
= 4 и
Результаты, полученные при численном исследовании задачи, отображены на рис. 4.8 и 4.9.
4.2. Исследование адекватности вариационной модели
Описание эмпирических данных, использованных при численной реализации модели. Эмпирические данные по динамике фитопланктонных сообществ, использованные для оценки и калибровки параметров модели и при ее верификации, были получены в ряде лабораторных экспериментов по культивированию планктонных водорослей (гл. 3).
С помощью данных, полученных в результате культивирования монокультур, рассчитывались начальные и минимальные клеточные квоты (по известным значениям начальных и конечных численностей и концентраций биогенных элементов), которые затем использовались для оценки параметров модели (ур. (7) в 4.1). Значения конечных численностей различных видов водорослей в поликультуре были использованы в процессе калибровки и верификации модели.
В том случае, когда в одном опыте использовалось несколько сред с различным содержанием биогенных элементов, опыту на каждой среде присвоен свой индекс, поставленный через точку после номера опыта (например, опыт 9.1 и 9.2).
При проверке адекватности теоретико-категорной модели были использованы материалы опытов 1, 2, 3, 4, 6 и 7 (см. 2.3). В опыте №1 сообщество из 10 видов выращивалось на среде с 195 мг/л азота и 5.2 мг/л фосфора. В опыте №2 сообщество из 8 видов выращивалось на 3 средах с одинаковым содержанием биогенных элементов (100 мг/л азота и 6 мг/л фосфора). В опыте №3 сообщество из 4 видов — на 5 средах с разным количеством фосфора (в порядке увеличения номера среды — 1.2, 2.2, 3.2, 4.2, 5.2 мг/л) и одинаковым количеством азота (28.5 мг/л). В опыте №4 сообщество из 4 видов выращивалось на двух средах с содержанием азота и фосфора соответственно 10.5 и 3.5 мг/л, 50
и 2.5 мг/л. В опыте №6 сообщество из трех видов выращивалось на трех средах, содержание азота и фосфора в которых составляло соответственно 0 и 3, 30 и 3, 60 и 3.5 мг/л. В опыте №7 сообщество из 4 видов выращивалось на трех средах, с содержанием азота в каждой 36.8 мг/л, а фосфора (в порядке увеличения номера среды) — 0.54, 1.04 и 1.54 мг/л.Качественная проверка адекватности модели с помощью рангового критерия для однофакторного лимитирования.
Для случая однофакторного лимитирования (m = 1) экстремальная модель сводится к степенному многочлену: ![]() . (15)
. (15)
Наиболее очевидным способом проверки модели была бы подстановка эмпирических значений клеточных квот с последующим решением уравнения (4) и сравнением вычисленных численностей сообщества с эмпирическими. Однако экспериментальные значения квот оцениваются косвенным методом и включают в себя ошибки измерения численностей и концентраций биогенов, в результате чего ошибка экспериментальной оценки клеточных квот может оказаться порядка 100%. Тем не менее существуют другие возможности проверки адекватности модели по результатам описанных выше экспериментов, например, сравнение ранговых структур сообществ из поликультур различных серий.
Под рангом вида мы понимаем порядковый номер вида в сообществе, ранжированном по численности. Из уравнения (15) следует, что ранг вида определяется рангом его клеточной квоты. В таком случае, ранговые структуры сообществ из поликультур различных серий должны определяться единой общей ранговой структурой квот. Таким образом, возможность нахождения единой ранговой структуры квот на качественном уровне будет свидетельствовать об адекватности модели.
При проверке адекватности модели с помощью рангового критерия были использованы эмпирические материалы серий опытов №1, 2 и 4 (клеточные квоты по азоту и фосфору, определенные в монокультурах данных серий, представлены в табл. 4.1 и 4.2). Сначала следует убедиться, что лимитирование в данной серии действительно происходит по одному фактору (так как в среде может иметь место лимитирование как по одному фактору, так и по обоим), и опытные результаты в принципе могут соответствовать модели (15).
Согласно модели в среде происходит лимитирование по одному фактору, если выполняется одно из условий (см. 4.3):
![]()
в этом случае лимитируют факторы D
1 или D2 соответственно. Если условие (16) не выполняется, то лимитируют оба фактора.Наибольшая сложность при проверке условия (16) состоит в том, что нам необходимо знать минимальные клеточные квоты. А они, как отмечалось выше, определяются из экспериментов в монокультурах со значительными ошибками. Для снижения влияния этих ошибок были составлены общие таблицы минимальных квот, определенных в монокультурах (табл. 4.1 и 4.2, см. также сводку квот в гл. 1), и для каждого вида водорослей выделены минимальные величины из набора минимальных квот. Именно с данными минимальными величинами производилась в дальнейшем проверка условия (16).
Обратимся к результатам экспериментов. В опытах №1 и 2 к моменту полного потребления фосфора азот оставался в среде, поэтому можно сделать вывод, что в данных сериях имело место лимитирование по фосфору. Опыт №4 проводился на двух средах. Проверим для них условие (2), причем в качестве минимальных квот для видов водорослей из опыта №4 возьмем абсолютные минимумы квот по данному виду из табл. 4.1 и 4.2. Результаты расчетов (табл. 4.3) показывают, что в опыте №4 на среде 1 наблюдалось двухфакторное лимитирование, а на среде 2 — однофакторное лимитирование по фосфору. В опыте №6 эксперимент проводился на трех средах. В среде 1 азота не было, поэтому можно считать, что лимитировал именно азот. Проверка условия (5) для второй и третьей сред показала, что на них имело место лимитирование по обоим факторам. Таким образом, эмпирической основой для построения модели выбраны результаты опытов 1, 2.1, 2.2, 2.3 и 4.2.
Процедура нахождения ранговой структуры квот состояла в переборе ее различных вариантов, сравнении по критерию Спирмена предсказываемых ранговых структур сообществ с экспериментальными (для всех имеющихся поликультур) и в выборе варианта, дающего максимальное соответствие. В результате был получен вариант ранговой структуры квот (табл. 4.4), который позволил с приемлемой точностью предсказать ранговую структуру сообществ всех поликультур: ранговый коэффициент корреляции Спирмена структур экспериментальных и предсказанных сообществ составляет от 0.915 до 1.
Остановимся несколько подробнее на смысле полученных результатов. Как уже отмечалось выше, невозможность использования экспериментальных значений квот вызвана в первую очередь тем, что эти величины оцениваются в экспериментах со значительными погрешностями. Именно поэтому значения клеточных квот для одного вида, но полученные на монокультурах различных серий (табл. 4.1 и 4.2), могут существенно различаться. Однако оказывается возможным найти такую единую ранговую структуру квот, на основе которой возможно адекватно описать ранговую структуру поликультур всех выбранных серий. Это может свидетельствовать, с одной стороны, о действительном постоянстве истинных значений минимальных клеточных квот, и, с другой стороны, об адекватности исходной формулы видовой структуры (4.1).
Количественная проверка адекватности при помощи калибровки параметров модели. Численная реализация математической модели включает в себя нахождение количественных значений параметров модели — ее калибровку, называемую также идентификацией или параметризацией. Для этого выбираются данные экспериментов или натурных наблюдений, характеризующие некие типичные ситуации, и по этим данным тестируются различные наборы параметров. Те из них, которые дают наиболее удачное согласие между модельными значениями выходных переменных и их экспериментальными значениями, используются в дальнейшем как основные наборы параметров модели.
В общем случае необходимость калибровки математических моделей экологических процессов объясняется несколькими причинами:
1. Данные, получаемые в результате экспериментальных исследований или натурных наблюдений, включают различные ошибки измерений. Модель, основанная на них, может давать результаты, отличные от реальных, именно за счет ошибок в определении исходных данных.
2. Для многих экологических параметров неизвестны их точные значения, а задаются лишь интервалы значений, поэтому имеет место неопределенность при выборе конкретного значения параметра модели.
3. Экологические модели являются упрощением реальности. Абсолютно все структурные связи исходной системы не могут быть включены в модель прямым образом. Если модель создается с достаточной тщательностью, то она описывает наиболее важные процессы и компоненты, но неучтенные детали все равно будут оказывать влияние на конечный результат. Возмущения такого рода могут быть косвенно учтены с помощью калибровки. Откалиброванные параметры могут иметь значения, несколько отличные от реальных, причем данное различие может быть обусловлено именно влиянием процессов, не включенных в модель.
Очевидно, что даже при самой тщательной калибровке результаты моделирования не могут абсолютно соответствовать реальным эмпирическим данным. Поэтому при работе с конкретной моделью выбирают определенный критерий калибровки, то есть критерий, по которому определяют допустимость различий между модельными и эмпирическими данными. Если критерий калибровки выполняется, то имеющийся набор параметров модели оставляют в качестве каркасного набора. Следует также отметить, что у одной модели может быть несколько наборов параметров, удовлетворяющих критерию калибровки.
Проверка качества модели обязательно должна включать в себя испытание откалиброванной модели на независимых, не использовавшихся при калибровке данных. Этот процесс испытания модели обозначается в литературе терминами “верификация” и “валидация”. Критерии верификации часто совпадают с критериями калибровки.
В случае неудачной верификации, то есть если модель с имеющимися наборами параметров плохо описывает эмпирические данные, не использовавшиеся в калибровке, зачастую приходится повторять поиск удачных наборов параметров при пересмотренных условиях калибровки (например, при изменении границ поиска параметров). Таким образом, разработка модели может быть итеративным процессом: процедуры калибровки и верификации приходится производить неоднократно.
В нашем случае основную сложность для проверки адекватности модели представляют ошибки в экспериментальной оценке значений минимальных клеточных квот. Однако существует возможность уточнения значений квот с помощью конечных численностей видов в поликультуре. Таким образом, задача калибровки состоит в нахождении такого набора клеточных квот, чтобы разность между реальными относительными численностями видов
j1 и предсказанными моделью относительными численностямиРассмотрим алгоритм калибровки для случая двухфакторного лимитирования, когда динамика сообщества описывается системой:
 (17)
(17)
Нам необходимо добиться примерного равенства:
j
1 ~Если взять в качестве начального приближения экспериментальные значения клеточных квот и решить с ними систему (17), то можно оценить численные значения x и y. Предположим, что
![]() .
.
Теперь решим систему (17) с новой квотой
![]() .
.
Для i-го вида мы несколько приблизились к верному значению квот.
Выполняем те же операции для всех остальных видов и повторяем процесс до тех пор, пока не будет выполнено условие:
 . (18)
. (18)
Описанную выше процедуру выполняем для всех имеющихся сред в экспериментальной серии. Для каждой среды мы получаем некоторый набор квот, решение же системы (17) с этими квотами получаем для всех имеющихся сред. После получения наборов квот по всем средам мы можем выбрать набор квот, наиболее адекватно описывающий динамику сообществ на всех средах, и считать калибровку законченной.
Легко заметить, что данный алгоритм совмещает процедуры калибровки и верификации, так как квоты подбираются только для одной среды, а проверяются по всему набору сред. Поэтому квоты, подобранные данным способом, можно считать верными, а модель (в случае удачного подбора квот) — адекватной.
Для однофакторного лимитирования алгоритм калибровки существенно упрощается, так как решать следует не систему (17), а уравнение (15). Вначале с помощью начальных приближений квот решается уравнение и определяется значение x. Дальнейшая процедура состоит в изменении значений квот, решении уравнения (7) из (4.1) и проверке условия (18) способом, аналогичным для двухфакторного лимитирования.
В качестве эмпирической основы для калибровки модели (6) были использованы результаты опытов №3 и 7. В опыте №3 сообщество из 4 видов (
Scenedesmus quadricauda, Scenedesmus obliquus, Ankistrodesmus sp., Chlorella vulgaris) выращивалось на 5 средах с разным количеством фосфора (в порядке увеличения номера среды — 1, 2, 3, 4, 5 мг/л) и одинаковым количеством азота (30 мг/л). Для всех 5 сред данной серии соотношения и абсолютные значения численностей видов в поликультуре были близки. Данный факт позволяет сделать вывод, что изменение концентрации фосфора в среде не влияло на развитие сообщества, а единственным лимитирующим фактором был азот. Следовательно, по материалам опыта № 3 можно откалибровать значения квот по азоту, используя алгоритм для однофакторного лимитирования.Результаты калибровки опыта №3 приведены в табл. 4.5. Для данного и остальных опытов в таблицах приводятся откалиброванные значения квот, экспериментальные и предсказанные значения абсолютных численностей и погрешность предсказания абсолютных численностей
D, вычисляемая по фор муле![]() .
.
При калибровке опыта №3 погрешность предсказания абсолютных численностей изменяется для разных видов от 3 до 49% и, как правило, укладывается в ошибку экспериментального определения численности.
В опыте №7 сообщество из 4 видов (
Scenedesmus quadricauda, Scenedesmus obliquus, Ankistrodesmus falcatus, Chlorella vulgaris) выращивалось на 3 средах, с содержанием азота во всех средах 30 мг/л, а фосфора (в порядке увеличения номера среды) — 0.5, 1 и 1.5 мг/л. Характер изменения абсолютных численностей у видов водорослей в поликультуре по разным средам различен. У одних видов (Chlorella vulgaris) изменения численности по средам практически отсутствуют (лимитирование по азоту), у других (Scenedesmus quadricauda, Scenedesmus obliquus) численность увеличивается с номером среды (лимитирование по фосфору), таким образом, можно предположить, что сообщество в целом лимитируется обоими факторами. Результаты калибровки опыта №7 приведены в табл. 4.6. Относительная погрешность предсказания абсолютных численностей составляет по разным видам от 6 до 78%.С помощью полученных значений минимальных клеточных квот по азоту и фосфору можно по условию (5) проверить правильность заключений о характере лимитирования в опыте №7, сделанных на основе анализа динамики численностей видов на различных средах. Результаты проверки (табл. 4.7) свидетельствуют о том, что на всех средах в опыте №7 действительно имело место двухфакторное лимитирование.
В результате калибровки параметров теоретико-категорной модели были получены наборы клеточных квот по азоту и фосфору, которые приемлемо описывают динамику сообщества на всех средах в пределах одного опыта. Это свидетельствует об адекватном отражении моделью динамики фитопланктонного сообщества в пределах одного опыта на количественном уровне.
Более общие выводы о количественной адекватности модели можно сделать на основе сопоставления значений минимальных клеточных квот, откалиброванных по различным опытам. Такое сравнение можно провести для квот по азоту из опытов №3 и 7 (для трех общих видов). При этом следует учесть, что уравнения экстремальной модели (7) из (4.1) инвариантны к преобразованию подобия квот
![]() ,
,
останутся прежними (поскольку вместе с параметрами системы алгебраических уравнений — квотами
Данное замечание не влияет на результаты проверки характера лимитирования, так как анализу подвергается именно соотношение квот. Однако для проведения сравнения значений квот между опытами разумнее оперировать отношениями квот к некоторой эталонной квоте (ею была выбрана квота
Scenedesmus quadricauda). Таким образом, соотношение калиброванных значений минимальных квот по азоту у видов Scenedesmus quadricauda, Scenedesmus obliquus, Chlorella vulgaris составляет соответственно в опыте №3 1:0.38:1.21, а в опыте №7 — 1:0.36:1.39, что свидетельствует о возможности описания с помощью схожих наборов параметров независимых наборов исходных данных.Существует также возможность сравнения рангов квот, найденных в опытах №1, 2 и 4 (табл. 4.4) с рангами калиброванных квот по фосфору из опыта №7 (табл. 4.6). Приемлемое соответствие рангов при таком сравнении отсутствует: ранги квот в табл. 4.4 у
Scenedesmus quadricauda, Scenedesmus obliquus, Ankistrodesmus falcatus, Chlorella vulgaris составляют соответственно 4, 2, 1, 3, а в табл. 4.6 — 2, 1, 4, 3. Следует отметить, однако, что основной вклад в различие ранговых структур вносит Ankistrodesmus falcatus. Данный вид на среде 2 опыта №7 имел значительное повышение численности, не интерпретируемое в рамках принятой модели, которое могло существенно повлиять на точность калибровки клеточных квот.Таким образом, в результате проведенных исследований можно сделать вывод о возможности нахождения набора минимальных клеточных квот, общего для всех опытов, и следовательно, об адекватности предсказания теоретико-категорной моделью (7) из (4.1) численностей лабораторных фитопланктонных сообществ.